Цитохром p450 характеристика та біологічна роль. P450 цитохроми
Полуніна Т.Є.
Оксана Михайлівна Драпкіна
– Ми продовжуємо нашу програму. У нас лекції та дискусії з гінекології закінчуються, ми повністю увійшли до регламенту, тому постараємося з нього не виходити. Професор Полуніна Тетяна Євгенівна відкриває секцію гастроентерології. Лекції «Роль сімейства цитохромів P450 у патогенезі та лікуванні неалкогольної жирової хвороби печінки».
Тетяна Євгенівна Полуніна, професор, доктор медичних наук:
- Цитохром P450 (CYP 450) - так називається велика родина універсальних ферментів організму людини. Цитохроми P450 відіграють важливу роль в окисленні численних сполук, таких як ендогенні сполуки (стероїди, жовчні кислоти, жирні кислоти, простагландини, лейкотрієни, біогенні аміни), а також екзогенні сполуки (лікарські препарати, продукти промислового забруднення, кантиди, пестициди) останні називаються ксенобіотиками.
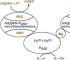
У цьому слайді ви бачите, де розташовані цитохроми P450. Вони розташовуються у гепатоциті, у цитозолі. Ендоплазматичний ретикулум є основою розташування. І, зокрема, ліпідна мембрана, що містить двошаровий шар фосфоліпідів, має кілька сполучених структур. Це цитохром, який включає в себе залізопротеїн, нікотинамідаденіндінуклеотид і оксидоредуктазу, які включаться в комплекс метаболізму лікарських препаратів і вище представлених ксенобіотиків.
Найбільш поширеними представниками цієї групи, до яких звертаються клініцисти, є цитохром P452 АС, P450 2D, P450 2E1, P450 3A4. Ці ферменти каталізують широкий спектр метаболічних реакцій та один цитохром, може метаболізувати декілька лікарських препаратів, що мають різну хімічну структуру. Один і той же лікарський препарат піддається різному впливу в цитохром P450 і в різних органах. І ось, зокрема, найважливіший цитохром, який ми звертаємо увагу, це цитохром P450 2E – найважливіший ізофермент цитохрому P450, він розщеплює ліпопротеїни низької щільності.
В даний час розроблені методи не тільки фенотипування, які ґрунтуються на субстратній специфічності певних ізоферментів цитохрому P450, але й активність того чи іншого ферменту та метаболізму визначається за фармакокінетикою маркерного субстрату та зміною концентрацій незміненої речовини та її метаболіту. А ось визначення ізоферментів цитохрому P450 шляхом ідентифікації відповідних генів ізоферментів здійснюється за допомогою полімеразної ланцюгової реакції. Це називається генотипуванням ізоферменту цитохрому P450.
На даному слайді ми бачимо, що в гепатоциті те місце, де розташовані ендоплазматичний ретикулум, цитохроми P450, яких налічується більше 50, і ліки, які розщеплюються в певному цитохромі, воно в ряді випадків з'єднується з цитохромом і утворює везикулу, яка пов'язує при цьому стрес та цитокіни; призводить до активізації туморнекротичного фактора і, зокрема, є тригерним фактором для запуску каспаз, що проявляється з каталітичних процесів.
Неалкогольна жирова хвороба печінки, яку згодом виділили в нозологічну одиницю, стали називати як неалкогольна жирова хвороба печінки (НАЖБП) з 1980 року, виявивши в печінці пацієнтів, що не зловживають алкоголем, зміни, які подібні до таких же, як при алкогольному ураженні.
Природний перебіг неалкогольної жирової хвороби печінки включає стеатоз як початкову стадію, яка, не прогресуючи, може протікати безсимптомно, і стеатогепатит, який супроводжується жахливими вегетативними проявами, синдромом цитолізу та диспепсичними проявами. При розвитку фіброзу виникає досить серйозна проблема – цироз печінки, та розвивається надалі портальна гіпертензія та карцинома.
Хотілося б звернути вашу увагу, що ще 1894 року Кірнан запропонував певну архітектоніку печінки, яка складається з балкової будівлі. На периферії балок, які складаються з гепатоцитів полігональної форми, розташована тріада: жовчний перебіг, портальна вена та артерія. Даний слайд представляє нормальну здорову печінку та жирову інфільтрацію гепатоцитів. Стеатоз печінки, який є однією з перших фаз розвитку неалкогольної жирової хвороби печінки, представлений у морфологічному вигляді на цій схемі.
Наступним варіантом розвитку запального процесу, який призводить до фіброзної тканини, поширення її печінки, ми бачимо стеатогепатит і надалі цироз печінки з розвитком портальної гіпертензії. Найчастіше це мікронодулярний цироз печінки, який вже цілком чітко затверджений у стадіях розвитку неалкогольної жирової хвороби печінки, він супроводжується портальною гіпертензією, варикозним розширенням вен стравоходу, шлунка, ускладненнями, які типові для цирозу печінки, та летальним кінцем.
При неалкогольному стеатогепатит найбільш часто розвиваються моменти, які пов'язані найчастіше як супутні захворювання: цукровий діабет, ожиріння. У пацієнтів неалкогольний стеатогепатит розвивається до 75%, а якщо поєднується цукровий діабет та ожиріння, то це вже 90% пацієнтів має неалкогольну жирову хворобу печінки.
Печінка, безсумнівно, головний орган-мішень, що уражується при метаболічному синдромі. Інсулінорезистентність – ключова ознака, яка є основою для накопичення ліпідів усередині гепатоцитів, жирової дистрофії печінки, неалкогольного стеатогепатиту та цирозу печінки.
Хотілося б звернути увагу, що метаболічний синдром включає не тільки порушення толерантності до глюкози, але й дисліпідемію, абдомінально-вісцеральне ожиріння, інсулінорезистентність та гіперінсулінемію, артеріальну гіпертензію, ранній атеросклероз, порушення гемостазу, гіперурикемію, гіперандроген. Хотілося б сказати, що неалкогольна жирова хвороба печінки, стеатоз, входить до складу метаболічного синдрому і є нині квінтетом, який раніше мав назву «смертельний квартет».
Фактори ризику, представлені на даному слайді, іноді змінюються у різних країнах, зокрема американські позиції та європейські позиції незначно відрізняються. Проте коло талії, рівень тригліцеридів, ліпопротеїдів, артеріальний тиск, зокрема 130/85, рівень глюкози – це ті показники, які необхідно обов'язково відстежувати у пацієнта з метаболічним синдромом.
Захворювання, асоційовані з ліпідним обміном, це неалкогольна жирова хвороба печінки, цукровий діабет другого типу, ішемічна хвороба печінки, гіпертонічна хвороба.
У схемі патогенезу особливе значення має інсулінорезистентність жирової тканини. Підвищення ліпогенезу, тобто збільшення рівня жирних кислот, збільшення синтезу тригліцеридів та ліпотоксичність призводять до того, що розвивається інсулінорезистентність, а це призводить до метаболічних дисфункцій, стресу ендоплазматичного ретикулуму, в якому також відбувається метаболізм жирних кислот та зокрема ліпопротеїдів, . Це клітини Купфера та зірчасті клітини, які надалі призводять не тільки до того, що підвищується рівень ліпідів дуже низької щільності, безсумнівно, це призводить до того, що розвивається стеатогепатит із фіброзом, і ми отримуємо активність процесу, що рухається у бік цирозу печінки.
На рівні гепатоциту жирні кислоти, які піддаються естерифікації в тригліцериди та експортуються у вигляді ліпопротеїдів низької щільності, це у нормальному гепатоциті ситуація, яка пов'язана з окисленням у мітохондріях, пероксисомах та мікросомах.
Безсумнівно, в механізмі інсулінорезистентності, який тут представлений, ключова роль належить туморнекротичному фактору, вільним радикалам, лептину, жирним кислотам і збільшення ліполізу, який призводить до поглинання жирних кислот, до порушення β-окислення жирних кислот у мітохондріях і також до гепатоцит.
Індукція цитохромів P450 4A11 і P450 2E1 призводить до перекисного окислення ліпідів, що призводить, безсумнівно, до активізації моментів, пов'язаних з накопиченням тригліцеридів. Гіперінсулінемія – ключовий момент, що веде до інсулінорезистентності. Він призводить також до збільшення гліколізу, синтезу жирних кислот та накопичення тригліцеридів у гепатоцитах.
У наступному слайді видно механізм взаємодії між мікросомальним окисленням та мітохондріальним β-окисленням. Зверніть увагу, що мітохондріальна Ω-оксидація та мітохондріальна β-оксидація призводять до запуску так званих рецепторів пероксисомального β-окислення і, зокрема, рецепторів, що активуються в проліферації пероксисом. Це призводить до експресії накопичення певного білка і, відповідно, ацетил-коензим А, який накопичується та запускає механізм, призводить до навантаження дикарбоновими жирними кислотами.
У наступному слайді ви бачите, що стеатогепатит та фіброз формуються на тлі активних мітохондріальних форм кисню. Ключовим моментом для запуску фіброзу є, поза сумнівом, накопичення малондіальдегіду, який призводить до формування запальних інфільтратів, фіброзу та активації зірчастих клітин. Зірчасті клітини запускають індукцію цитокінів, таких як туморнекротичний фактор, що трансформують фактори росту. Виснаження антиоксидантної системи призводить до запуску Fas-леганду, мітохондріальної активної форми кисню, виникає некроз гепатоциту, і надалі розвивається фіброзна тканина, яка є основою для розвитку цирозу.
На даному слайді представлена схема, ви бачите надлишок ліпідів, які накопичуються у гепатоциті. Мітохондріальна дисфункція та дисфункція цитохрому P450 призводить до активізації перекисного окислення ліпіду, запуску клітини Купфера, запальних цитокінів, активації зірчастих клітин та апоптозу, який надалі призводить до розвитку некрозу гепатоциту.
Метаболічний синдром дуже важливий, тому що неалкогольна жирова хвороба печінки – це частина метаболічного синдрому. І не тільки на гепатоцит, в якому відбувається підвищення рівня ліпопротеїдів низької щільності та дуже низької щільності, тригліцеридів (це дуже важливо), але також впливає на ендотеліальну клітину. Відбувається ендотеліальна дисфункція та запускається також момент, який пов'язаний із перекисним окисленням ліпідів, накопиченням речовин, що впливають на атеросклероз, раптову смерть, серцеві напади.
Безперечно, підвищення рівня вільних жирних кислот пов'язане з адипоцитами. І зниження зокрема естерифікованого холестерину також призводить до різних стресів ядерного рецептора. А особливо важливий нині так званий активований рецептор проліферації пероксисом, саме до нього спрямовані всі погляди вчених, які працюють з ожирінням, діабетом, неалкогольною жировою хворобою печінки.
Моноцит (макрофаг) у ряді випадків підвищенням рівня запальних відповідачів (туморнекротичного фактора, інтерлейкінів-6, мембранні толл-подібні рецептори, вільні жирні кислоти) також запускає моменти, пов'язані саме з патологічними ефектами жирних кислот.
Критерії оцінки інсулінорезистентності всім відомі ще з 1985 року. Її визначають індекс HOMA – Homeostasis Model Assessment, і вже сучасніший індекс QUICKI – Quantitave Insulin Sensitivity. Тут представлено концентрацію інсуліну, глюкоза сироватки крові, а також норми.
Хотілося б звернути увагу на те, що не всім пацієнтам з неалкогольною жировою хворобою печінки необхідно проводити біопсію печінки. У нас зараз є моменти, які дають можливість нам визначити рівень жирової інфільтрації печінки. І, зокрема, це фібротест.
В алгоритмі діагностики неалкогольної жирової хвороби печінки ми приділяємо увагу не лише специфічним ознакам, але також активності ферментів аланінової та аспарагінової трансамінази, гамма-глутамілтранспептидази, лужної фосфатази, звертаємо увагу на прийом алкоголю, який обговорювався у попередніх колег. І я хотіла б звернути увагу, звісно, на фактори ризику: метаболічний синдром, інсулінорезистентність, цукровий діабет. Призначається терапія корекції даної ситуації, за необхідності біопсія печінки. Безсумнівно, потрібні абсолютні показання щодо біопсії. І якщо індекс маси тіла перевищує 35 та 40, тоді вже проводяться заходи, пов'язані з хірургічним лікуванням.
Хотіла б звернути вашу увагу на ряд лікарських препаратів (нестероїдні – протизапальний глюкокортикоз, і стероїдні препарати, антибіотики тетрациклінового ряду), ряд нутриціологічних факторів (голодування, швидке зниження ваги, хірургічні втручання, метаболічні генетичні фактори, зокрема та інші супутні захворювання. Це дуже важливо задля диференціальної діагностики.
У стадії стеатозу важливим є лікування ожиріння, інсулінорезистентності, дисліпідемії. У стадії стеатогепатиту найважливішим моментом є усунення окислювального стресу, запалення та фіброзу.
Надмірна індукція цитохрому P450 2E шкідливо впливає на гепатоцити внаслідок вивільнення вільних радикалів. Есенційні фосфоліпіди діють не тільки як антиоксиданти, але також є дуже важливим моментом для зниження активності цитохрому 2E1, що показано в роботах М. Алейник. Результати деяких досліджень свідчать, що введення есенціальних фосфоліпідів може зменшити індукцію цитохрому P450 2E (роботи Володимира Трохимовича Івашкіна, які були представлені з Мариною Вікторівною Маєвською у російських джерелах у 2004 році).
У формуванні кінцевої стадії неалкогольної жирової хвороби печінки беруть участь стелатні клітини. І в лабораторних експериментах було продемонстровано, що повне запобігання активації стеллатних клітин із застосуванням інгібіторів CYP2E1 запобігають розвитку цирозу.
Хотіла б звернути вашу увагу, що не тільки російський автор М.Алійник, а також японський автор Акіяма в журналі «Гепатологія» у 2009 році на підставі моделі алкогольного ураження печінки також приділяє увагу цитохрому P450 2E, ацетил-КоА оксидазе та нікотинамідаденіді що есенціальні фосфоліпіди виявляють протизапальну антиапоптотичну та протифіброзну активність при даній патології.
Це теоретичний варіант припущення застосування інгібіторів цитохромів P450, зокрема препарату «Есенціалі», який є еталонним, і є найважливішим моментом для інгібування цитохромів P450 2E і, відповідно, P450 4A11. Це запобігає окисленню ліпідів, гліколізу та знижує синтез жирних кислот.
У лікуванні неалкогольної жирової хвороби печінки представлені препарати: інсулінові сенситайзери, антиоксиданти, гепатопротектори, антимікробні препарати.
Але я хотіла б звернути увагу на фосфоліпіди мембран. Вони є основними ліпідними компонентами клітинних мембран. Ушкодження фосфоліпідних мембран веде до синдрому цитолізу, а надлишок реактивних складових кисню призводить до пошкодження фосфоліпідних мембран на підставі мікросомального γ-окислення та пероксимального β-окислення. Відповідно, пошкодження фосфоліпідних мембран – це загибель клітини, що призводить до запуску фіброзу та активації зірчастих клітин.
Ушкодження структури печінки – це ушкодження мембран. У варіанті есенціальних фосфоліпідів є матеріал, який відновлює клітинні мембрани замість ліпідів. Відновлення структури печінки дає можливість відновлення функції печінки.
Наші пацієнти страждають не тільки на алкогольну жирову хворобу печінки, алкогольні гепатити, а є й інші захворювання печінки, це факт незаперечний. Хотіла б звернути вашу увагу, що за даними Е. Кунца (монографія 2008 року) есенційні фосфоліпіди мають антифібротичний ефект, дію, що стабілізує жовч і мембрану гепатоциту.
Це видання, яке було випущено у 2008 році на підставі фармакологічних та клінічних даних. Терапія есенціальними фосфоліпідами є кращим вибором для значного зменшення прояву та усунення жирової хвороби печінки різної етіології, що розвинулася внаслідок вживання алкоголю, ожиріння і навіть причину не вдається побачити.
Я хотіла б звернути увагу на те, що існує кілька досліджень з «Есенціалі». Ці дослідження досить відомі. Але хотіла б сказати, що і з цукровим діабетом «Есенціалі» дає можливість у пацієнтів із неалкогольною хворобою печінки нормалізувати рівень глюкози, глікованого гемоглобіну, сироваткового холестерину.
І на закінчення хотіла б сказати, що ураження печінки, що характеризується накопиченням жиру за відсутності зловживання алкоголем, відомо як неалкогольна жирова хвороба печінки. Чинниками ризику є ожиріння, цукровий діабет другого типу. У патогенезі неалкогольної жирової хвороби печінки особливе значення приділяється надмірній активності цитохромів P450 2E1. Клінічні варіанти перебігу хвороби: біль у правому підребер'ї, астеновегетативні та диспепсичні розлади, гепатомегалія. І наш алгоритм діагностики заснований на послідовному виключенні алкогольних та ятрогенних, а також вірусних уражень печінки.
Мікросомальне окислення – це послідовність реакцій за участю оксигеназі НАДФН, що призводять до впровадження атома кисню до складу неполярної молекули і появі у неї гідрофільності і підвищує її реакційну здатність.
Реакції мікросомального окисненняздійснюються кількома ферментами, розташованими на мембранах ендоплазматичного ретикулуму (у разі in vitroвони називаються мікросомальні мембрани). Ферменти організують короткі ланцюги, які закінчуються цитохромом P 450 .
Реакції мікросомального окиснення відносяться до реакцій фази 1і призначені для надання гідрофобної молекули полярних властивостей та/або підвищення її гідрофільності, посилення реакційної здатності молекул для участі в реакціях 2 фази. У реакціях окислення відбувається утворення або вивільнення гідроксильних, карбоксильних, тіолових та аміногруп, які є гідрофільними.
Ферменти мікросомального окислення розташовуються в гладкому ендоплазматичному ретикулумі і є оксидазами зі змішаною функцією(Монооксигеназ).
Цитохром P450
Основним білком мікросомального окиснення є гемопротеїн - цитохром Р 450 .У природі існує до 150 ізоформ цього білка, що окислюють близько 3000 різних субстратів. Співвідношення різних ізоформ цитохрому Р450 відрізняється з генетичних особливостей. Вважається, що одні ізоформи беруть участь у біотрансформації ксенобіотиків, інші метаболізують ендогенні сполуки (стероїдні гормони, простагландини, жирні кислоти та ін.).
Цитохром Р450взаємодіє з молекулярним киснем і включає один атом кисню в молекулу субстрату, сприяючи появі (посилення) у неї гідрофільності, а інший – молекулу води. Основними його реакціями є:
- окисне деалкілування, що супроводжується окисленням алкільної групи (при атомах N, O або S) до альдегідної та її відщепленням,
- окислення (гідроксилювання) неполярних сполук з аліфатичними або ароматичними кільцями,
- окиснення спиртів до відповідних альдегідів.
Робота цитохрому Р 450 забезпечується двома ферментами:
- НАДН-цитохром b 5 -оксидоредуктаза, містить ФАД,
- НАДФН-цитохром Р 450-оксидоредуктаза, містить ФМНі ФАД.

Схема взаєморозташування ферментів мікросомального окиснення та їх функції
Обидві оксидоредуктази одержують електрони від відповідних відновлених еквівалентів та передають їх на цитохром Р 450 . Цей білок, попередньо приєднавши молекулу відновленого субстрату, зв'язується із молекулою кисню. Отримавши ще один електрон, цитохром P 450 здійснює включення до складу гідрофобного субстрату першого кисню атома (окислення субстрату). Одночасно відбувається відновлення другого атома кисню до води.

Послідовність реакцій гідроксилювання субстратів за участю цитохрому Р450
Істотною особливістю мікросомального окислення є здатність до індукції чи інгібування, тобто. до зміни потужності процесу.
Індукторами є речовини, що активують синтез цитохрому Р 450 та транскрипцію відповідних мРНК. Вони бувають
1. Широкого спектрудії, які мають здатність стимулювати синтез цитохрому Р 450 , НАДФН-цитохром Р 450 -оксидоредуктази та глюкуронілтрансферази. Класичним представником є похідні барбітурової кислоти. барбітурати,також до цієї групи входять діазепам, карбамазепін, рифампіцинта ін.
2. Вузький спектра події, тобто. стимулюють одну з форм цитохрому Р 450 – ароматичні поліциклічні вуглеводні ( метилхолантрен, спіронолактон), етанол.
Наприклад, етанолстимулює синтез ізоформи Р 450 2Е1 (алкогольоксидаза) яка бере участь у метаболізмі, етанолі, нітрозамінах, парацетамолу та ін.
Глюкокортикоїдиіндукують ізоформу Р 450 3А.
Інгібітори мікросомального окиснення зв'язуються з білковою частиною цитохрому або із залізом гему. Вони поділяються на:
1. Оборотні
- прямогодії- чадний газ ( СО), антиоксиданти,
- непрямогодії, тобто. впливають через проміжні продукти свого метаболізму, які утворюють комплекси із цитохромом Р 450 – еритроміцин.
2. Необоротніінгібітори – алопурінол, аміназин, прогестерон, оральні контрацептиви, тетурам, фторурацил,
Оцінка реакцій 1-ї фази
Оцінку мікросомального окиснення можна проводити такими способами:
- визначення активності мікросомальних ферментів після біопсії,
- з фармакокінетики препаратів,
- за допомогою метаболічних маркерів ( антипіринова проба).
Антипіринова проба
Обстежуваний приймає вранці натще. амідопіриніз розрахунку 6 мг/кг ваги. Збирається 4 порції сечі в інтервалі відповідно від 1 до 6 годин, 6-12, 12-24 та 45-48 годин. Об'єм сечі вимірюється. Не пізніше ніж через 24 години сеча центрифугується або фільтрується. Далі досліджується концентрація 4-аміноантипірину та його метаболіту N-ацетил-4-аміноантипірину в сечі.
Цитохром Р450
Суперсімейство цитохромів Р-450 (CYP-450) відповідає за мікросомальне окислення і є групою ферментів, що мають безліч ізоформ (більше 1000), які не тільки здійснюють метаболізм ліків, але й беруть участь у синтезі стероїдних гормонів, холестерину та інших речовин.
Найбільша кількість цитохромів виявлена в гепатоцитах, а також у таких органах, як кишечник, нирки, легені, головний мозок, серце. Ізоферменти цитохромів на підставі гомології нуклеотидної та амінокислотної послідовностей поділяють на сімейства, які, у свою чергу, поділяють на підродини. Представники різних сімейств відрізняються субстратною специфічністю та регуляторами активності (індуктори та інгібітори). Хоча окремі члени сімейств можуть мати «перехресну» специфічність та «перехресні» індуктори та інгібітори. Так, показано, що противірусний препарат ритонавір метаболізується сімома ферментами (CYP1A1, CYP2A6, CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4), а циметидин інгібує чотири ферменти (CYP1A2, CY Найбільш важливими для біотрансформації ліків є цитохроми CYP1A1, CYP2A2, CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4, CYP3A5. Відносний внесок різних цитохромів та інших ферментів І фази детоксикації у метаболізмі ліків представлений малюнку 7.2.2.
Кожен ізофермент цитохрому Р-450 кодується своїм геном, який локалізується на різних хромосомах. Частина таких генів має близько розташовані до них псевдогени (копії, що не експресуються), які істотно ускладнюють проведення генетичного тестування.
Внаслідок поліморфізму генів метаболізму активність відповідних ферментів у різних осіб може суттєво варіювати. Залежно від цих міжіндивідуальних особливостей виділяють три групи осіб, які різняться за активністю того чи іншого ферменту метаболізму. Це так звані «екстенсивні» метаболізатори – особи з нормальною швидкістю метаболізму ліків (основна частина популяції), «повільні» метаболізатори (особи зі зниженою швидкістю метаболізму певних ліків) та «швидкі» («надактивні») метаболізатори – індивіди з підвищеною швидкістю біотрансформація деяких ліків. Частка «повільних» і «швидких» метаболізаторів щодо окремих ферментів метаболізму виявляє суттєві міжпопуляційні відмінності. Водночас далеко не завжди відзначається повна кореляція генотипу та фенотипу у швидкості метаболізму ліків, що свідчить про необхідність використання біохімічного контролю при генотипуванні ферментів метаболізму.
Розглянемо функціональні особливості поліморфізму основних генів суперродин цитохромів CYP-450, що беруть участь у метаболізмі ліків. Детальну інформацію про властивості ферментів метаболізму, їх субстратні характеристики та генетичний поліморфізм можна знайти в серії вітчизняних монографій та підручників з клінічної фармакогенетики.
Сімейство Р-450 CYP1 метаболізує порівняно невелику частину ксенобіотиків, найважливіші з яких є поліциклічними ароматичними вуглеводнями (ПАУ) - основними компонентами тютюнового диму.
Особливо важлива роль цьому належить генам CYP1A1 і CYP1A2, локалізованим на хромосомі 15. Експресію обох генів регулює комплекс, утворений Ah-рецептором з індукуючої молекулою ПАУ, який проникає в ядро і специфічно стимулює експресію цих генів.
CYP1A1 кодує білок з арилгідрокарбонат-гідроксилазною активністю, що контролює початковий метаболізм ПАУ, що призводить до утворення канцерогенів (наприклад, бензопірену, що утворюється при тютюнокурінні). Генний поліморфізм CYP1A1 обумовлений трьома точковими мутаціями: С4887А та A4889G в екзоні 7 та T6235C у 3'-фланкуючій ділянці. Заміна G4889 (Val) + C6235 характеризується появою "швидкого" алелю * 2B. Він має в 3 рази більш високу активність порівняно з алелем дикого типу. *2B зустрічається майже у 7% представників європеоїдної раси і розглядається як фактор ризику раку легень. Показано, що за наявності *2B-алелю у курців ризик розвитку раку легені порівняно з некурцями зростає більш ніж у сім разів. Ризик стає ще більшим, якщо, крім алелю *2B гена CYP1A1, індивід, що палить, має також «неповноцінний» алель гена GSTM1. Алелі *2A (C6235) і *4 (A4887(Asp) зустрічаються в популяції з частотою всього 1-3%. При цьому *2A-аллель асоційований зі спадковою схильністю до лейкозу та резистентністю до лікарської терапії цього захворювання.
Продукт гена CYP1A2 метаболізує на тільки ПАУ, але й такі сполуки, як кофеїн, теофілін та ін.
Сімейство Р-450 CYP2 - представлено групою функціонально найбільш значущих ферментів, що метаболізують величезну кількість різних препаратів. Їхня активність виявляє виражену залежність від генетичного поліморфізму.
Підродина CYP2A є найважливішим ізоферментом цієї підродини. Він бере участь у перетворенні нікотину на котинін, у гідроксилюванні кумарину та циклофосаміду, робить внесок у метаболізм ритонавіру, парацетамолу та вальпроєвої кислоти. CYP2A6 бере участь у біоактивації компонентів тютюнового диму - нітрозамінів, що викликають рак легень. Ген CYP1A6 локалізовано на 19 хромосомі в локусі 19q13.2. В основному ген експресується у печінці. Показано, що алель *4 гена CYP1A6 є протективним, тобто асоціюється з меншим ризиком виникнення раку легені. Наявність алелів *2 і *3 асоційовано зі зниженим метаболізмом кумарину, що має значення при дозуванні цього препарату через можливу гепатотоксичну дію.
Підродина CYP2B. Усі ферменти цієї підродини індукуються фенобарбіталом. Найбільш значущим ферментом є CYP2B6, який метаболізує багато цитостатиків (циклофосамід), противірусні препарати (ефавіренц і невірапін), антидепресанти (бупропіон), анестетики (пропофол) і снітетичні опіоїди (метадон), а також бере участь в. Ген CYP2B6 локалізований у тому ж локусі, що і ген CYP2A6, екпресується переважно в печінці. Наявність повільних алелей гена CYP2B6 (*2, *4, *5, *6) знижує швидкість метаболізму противірусних препаратів, що призводить до зниження кліренсу та підвищує ризик ускладнень з боку ЦНС.
Підродина CYP2C відіграє ключову роль у метаболізмі багатьох ліків. Загальною властивістю цих ізоферментів є наявність 4-гідролазної активності щодо протисудомного препарату мефенітоїну.
Особливо важливим для клінічної фармакогенетики є тестування поліморфізму гена CYP2C9, локалізованого у локусі 10q24. Ген екпресується переважно в печінці, є головним метаболізатором інгібіторів ангіотензинових рецепторів (лозартану та ірберсартану). Його субстратами також є антикоагулянти (варфарин), цукрові препарати (гліпізид), протисудомні препарати (фенітоїн, діазепам), антидепресанти (амітриптилін, кломіпрамін, іміпрамін), інгібітори протонових помп (омепразол), нестероїдні протиовос , Толбутамін . Як згадувалося, аналіз поліморфізму гена CYP2C9 став першим офіційно схваленим генетичним тестом (див. вище). Кількість індивідів, що мають знижену активність даного ферменту, у вітчизняній популяції становить до 20%. При цьому, щоб уникнути небажаних побічних ефектів, лікувальну дозу перерахованих вище препаратів у носіїв алелів *2 і *3 гена CYP2C9 необхідно зменшувати в 2-4 рази.
Ген CYP2C19 локалізований у локусі 10q24.1-q24.3, експресується у печінці. Його білковий продукт є основним ферментом метаболізму інгібіторів протонного насоса (омепразол) та протисудомних препаратів (прогуаніл, вальпроєва кислота, діазепам, барбітурати). Частота його «повільного» алелю (*2) у європейській популяції коливається від 5 до 20 0%.
Підродина CYP2D. Цитохром CYP2D6 метаболізує близько 20% усіх відомих лікарських засобів. Ген CYP2D6 локалізовано на 22 хромосомі в локусі 22q13.1. Основним місцем його експресії є печінка. В даний час у гені CYP2D6 ідентифіковано більше 36 алелів, деякі з них характеризуються відсутністю білкового продукту, інші призводять до появи ферменту зі зміненими властивостями. Субстратами ферменту CYP2D6 є такі широко використовуються в клінічній практиці лікарські засоби, як бета-адреноблокатори, антидепресанти, антипсихотропні речовини, антиаритмічні, нейролептики, протигіпертонічні препарати, інгібітори монооксидредуктази, похідні морфіну, нейротрансмітери (додатки). Зважаючи на те, що близько 6-10 % європеоїдів належать до повільних метаболізаторів цього ферменту, очевидна необхідність у генетичному тестуванні CYP2D6 з метою корекції доз згаданих препаратів. Крім того, «функціонально ослаблені» алелі цього гена асоційовані зі спадковою схильністю до таких тяжких хвороб, як рак легені, рак кишечника та ін.
Підродина CYP2E. Цитохром CYP2E1 відноситься до етаноліндуцибельних ферментів. Його субстратами є карбонтетрахлорид, диметилнітрозамін. Є дані про те, що CYP2E1, поряд з CYP1A2, бере участь у перетворенні парацетамолу в N-ацетилбензохінонімін, що має потужну гепатотоксичну дію. Крім того, він є найважливішим ізоферментом групи цитохромів, що окислюють холестерин ліпопротеїнів низької щільності, що, у свою чергу, веде до утворення атеросклеротичних бляшок. Ген CYP2E1 локалізований у локусі 10q24.3-qter, експресується у печінці дорослих людей. Taq1-поліморфізм у гені CYP2E1 призводить до зниження активності даного ферменту. Гомозиготи M/M за ослабленим алелем гена CYP2E1 виявляють підвищену чутливість до вищевказаних препаратів внаслідок їх уповільненої детоксикації.
Сімейство цитохрому Р-450 CYP3
Підродина CYP3A найбільш численна. На його частку припадає близько 30% усіх ізоферментів цитохрому Р-450 у печінці та 70% всіх ізоферментів стінки шлунково-кишкового тракту. Найбільш значущими є ферменти CYP3A4 та CYP3A5, гени яких локалізовані у локусі 7q22.1. У печінці екпресується переважно ген CYP3A4, а у шлунково-кишковому тракті – CYP3A5.
Фермент CYP3A4 метаболізує понад 60% усіх ліків і відіграє велику роль у метаболізмі тестостерону та естрогенів. Алельні варіанти гена CYP3A4 дуже численні, але дані про їх вплив на фармакокінетику відповідних лікарських засобів суперечливі.
Фермент CYP3A5 метаболізує частину ліків, із якими взаємодіє CYP3A4. Показано, що наявність алелю *3 гена CYP3A5 призводить до зниження кліренсу таких ліків, як альпразали, мідазоли, саквінавір.
Параоксоназа – фермент, що відповідає за синтез параоксонази – білка плазми крові. Крім цього, фермент інактивує фосфорорганічні сполуки, органофосфати, карбамати, ефіри оцтової ксилоти. Частина цих речовин є бойовими отруйними речовинами - зарин, зоман, табун. З відомих трьох ізоформ найбільше значення має фермент PON1. Ген його локалізовано у локусі 7q21.3. Найбільш значущим та вивченим поліморфізмом є заміна глютаміну на аргінін у 192 положенні (L/M-поліморфізм). Показано, що аллель М асоційований зі зниженим метаболізмом фосфорорганічних сполук.
Алель М та М/М-генотип збільшують ризик розвитку хвороби Паркінсона, особливо у поєднанні з 5-алелем гена GSTP1, та асоційовані з утворенням атеросклеротичних бляшок.
Алкоголь- та альдегіддегідрогенази
Алкогольдегідрогеназа є ключовим ферментом у катаболізмі етанолу та інших спиртів, окислюючи спирти до альдегідів. У дорослої людини ген ADH1B експресується у печінці. Існує певна динаміка рівня його експресії залежно віку. Ген ADH1B (ADH2) локалізований у локусі 4q22. Найбільш вивчений поліморфізм – G141A. Показано, що аллель А пов'язаний з підвищеною активністю ферменту, що призводить до надмірного накопичення проміжних продуктів метаболізму - альдегідів, що мають виражений токсичний ефект. Індивідууми з алелем гена ADH1B мають підвищену чутливість до етанолу і менш схильні до алкоголізму.
У клітинах печінки присутні також дві альдегіддегідрогенази: ALDH1 (цитозольна) та ALDH2 (мітохондріальна). Ген ALDH2 локалізований у локусі 12q24.2, його продукт відіграє ключову роль у перетворенні токсичних альдегідів у відповідні карбонові кислоти, які легко видаляються з організму. ALDH2 відіграє у катаболізмі алкоголю. Відомо, що представники жовтої раси алкогольна інтоксикація обумовлена відсутністю ALDH2 майже в 50 % населення. Поліморфізм в гені ALDH2 призводить до заміни Glu в положенні білка 487 (ALDH2*1-аллель) на Lys (ALDH2*2-аллель). ALDH2*2-аллель кодує фермент зі зниженою активністю. У гетерозигот активність ферменту знижена вдесятеро. Фермент ALDH2 залучений до патогенезу різних раків, пов'язаних із надмірним споживанням алкоголю, - гепатоцелюлярна карцинома, рак стравоходу, глотки та ротової порожнини.
Інтенсивний прийом алкоголю в осіб з несприятливими алельними варіантами генів ADH1B та ALDH2 може призвести до швидкого розвитку печінкових ускладнень: алкогольної хвороби та цирозу печінки.
Цитохром P450, сімейство 2, субсімейство C, поліпептид 9 (CYP2C9). Виявлення мутації A1075C (Ile359Leu)
Назва гена -CYP2C9
Локалізація гена на хромосомі- 10q23.33
- *1/*1
- *1/*3
- *3/*3
Зустрічаємось у популяції
Алель CYP2C9*3зустрічається у європейців із частотою 6 %.
Асоціація маркера з метаболізмом лікарських препаратів
Досліджується для виявлення фізіологічної ефективності застосування препаратів: пероральних антикоагулянтів із класу кумаринів (варфарин), похідних сульфонілсечовини, ненаркотичних анальгетиків (теноксикам, флурбіпрофен, лорноксикам, піроксикам), лозартану та ірбесартану (блок).
Загальна інформація про дослідження
Для запобігання та лікування тромбоемболічних ускладнень найчастіше застосовується препарат варфарин ("Кумадин"). Він призначається для тривалого прийому в серії випадків, пов'язаних з підвищеною згортанням крові, а також післяопераційний період з метою запобігання утворенню тромбів внаслідок хірургічного втручання. Часто практикується призначення препарату людям, які перенесли інсульти, інфаркт міокарда.
Для досягнення ефекту лікарських препаратів необхідна їхня біоактивація в організмі (трансформація в активну форму) у клітинах печінки (гепатоцитах) системою ферментів цитохрому Р450 (CYP). Гени, що кодують ці ферменти, поліморфні, причому часто зустрічаються алелі, що кодують утворення ферментів зі зниженою або відсутньою функцією.
На активність цитохромів, крім особливостей будови генів, що кодують, впливають такі фактори, як вік, маса тіла, спосіб життя, шкідливі звички, особливості дієти, супутні захворювання, прийом лікарських препаратів. Ці чинники відповідають формування індивідуальних особливостей роботи ферментів Р450 і визначають характер метаболізму більшості ліків. Головним ферментом біотрансформації непрямих антикоагулянтів є ізофермент цитохрому Р450 CYP2C9.
Ген CYP2C9локалізовано на 10-й хромосомі в ділянці 10q23.33. Існують варіанти гена (алелі) CYP2C9, що кодують утворення ферменту зі зниженою або відсутньою функцією Варіант гена, що несе точкову заміну аденіну на цитозин у позиції 1075 (A1075C), веде до зниження метаболічної активності ферменту та позначається як CYP2C9*3. Заміна одного нуклеотиду спричиняє заміну амінокислоти ізолейцину на лейцин (Ile359Leu) у ферменті CYP2C9. Таким чином, синтезується фермент із зміненою функцією, активність якого становить менше ніж 5 % від активності ферменту *1. Основний (незмінений) варіант гена позначається як CYP2C9*1.
Найбільш поширений генотип, що зумовлює нормальний метаболізм варфарину і позначається як CYP2C9*1/*1.
Генетичний маркер CYP2C9*3(генотипи *3/*3 та *3/*1) пов'язаний із зміною функціональної активності ферменту цитохрому P450, що знижує швидкість виведення варфарину з організму. Наявність у пацієнта алелю *3 призводить до суттєвого зниження активності ізоферменту цитохрому, що збільшує антикоагуляційний ефект препаратів до 7 разів і може спричиняти розвиток таких ускладнень, як великі внутрішні кровотечі та епізоди надмірної гіпокоагуляції.
Цитохром Р450. Структура та функція
Серед ферментів 1-ї фази чільне місце займає система цитохрому Р450 (P450 або CYP) з точки зору каталітичної активності щодо величезної кількості ксенобіотиків. Найбільша концентрація цитохрому Р450 виявляється у ендоплазматичному ретикулумі гепатоцитів (мікросомах). Печінкові мікросомальні цитохроми Р450 відіграють найважливішу роль у визначенні інтенсивності та часу дії чужорідних сполук та ключову - у детоксикації ксенобіотиків, а також в активації їх до токсичних та/або канцерогенних метаболітів. Цитохром Р450-залежні монооксигенази - мультиферментна електрон-транспортна система. Всі цитохроми Р450 - білки, що містять гему. Зазвичай гемове залізо перебуває у окисленому стані (Fe3+). Відновлюючись до Fe2+, цитохром Р450 здатний зв'язувати ліганди, такі як кисень або монооксид вуглецю. Комплекс відновленого цитохрому Р450 з З має максимум поглинання 450 nм, що і стало підставою для
назви цих ферментів. Основна реакція, яку каталізують цитохроми Р450 – монооксигеназна, в якій один атом кисню взаємодіє із субстратом (RH), а інший відновлюється до Н2О. Як відновник у реакції бере участь НАДФН:
RH (субстрат) + О2 + НАДФH + H + --> ROH (продукт) + Н2О + НАДФ +
Механізм, завдяки якому цитохром одержує електрон від НАДФH, залежить від внутрішньоклітинної локалізації цитохрому Р450. В ЕПР, де розташована більшість гемопротеїдів, що беруть участь у біотрансформації ксенобіотиків, електрон передається через флавопротеїн, що називається НАДФН-Р450 редуктазу. Одна молекула редуктази може доставляти електрони кілька різних молекул Р450. У мітохондріях, де розташовані ітохроми Р450, що беруть участь у біосинтезі стероїдних гормонів та метаболізмі вітаміну D, електрон переноситься за допомогою 2-х білків: ферродоксину або ферродоксин-редуктази.
На рис. 1 показаний каталітичний цикл цитохрому Р450. 1-а частина циклу полягає в активації кисню, 2-а – в окисненні субстрату. Схема дії мікросомальної монооксигеназної системи вперше була описана Естабруком з співавт., В даний час вона підтверджена багатьма дослідниками. Ця схема така: перша стадія полягає у взаємодії субстрату з окисленою формою Р450. При зв'язуванні Р450 із субстратами
відбувається перехід гемового заліза з низькоспінового у високоспиновий стан. Друга стадія полягає у відновленні фермент-субстратного комплексу, що утворився, першим електроном, який надходить з НАДФН-специфічного ланцюга переносу від НАДФН через
флавопротеїд I (НАДФН-цитохром Р450 редуктазу) Третя стадія полягає у освіті потрійного комплексу: відновлений цитохрому Р450-субстрат-кисень. Четверта стадія
являє собою відновлення потрійного комплексу другим електроном, який, як
вважають, надходить з НАДН-специфічного ланцюга перенесення електронів, що складається з НАДН-
цитохром b5 редуктази або флавопротеїду II та цитохрому b5. П'ята стадія складається з кількох процесів, що включають внутрішньомолекулярні перетворення відновленого потрійного комплексу та його розпад з утворенням гідроксильованого продукту та води. На цій стадії цитохром Р450 перетворюється на вихідну окислену форму.
 Цитохроми Р450 каталізують такі типи реакцій: гідроксилювання аліфатичного або ароматичного атома вуглецю; епоксидування подвійного зв'язку;
Цитохроми Р450 каталізують такі типи реакцій: гідроксилювання аліфатичного або ароматичного атома вуглецю; епоксидування подвійного зв'язку;
окислення атома (S, N, I) або N-гідроксилювання; перенесення окисленої групи;
руйнування ефірного зв'язку; дегідрогеніювання. Деякі реакції, що каталізуються
цитохромом Р450 представлені на рис. 2 та 3. Декілька класів реагентів добре
гідроксилюється останній вуглець у ланцюгу, так зване омега-гідроксилування. Так
А буває внутрішнє гідроксилювання на кількох позиціях (позиції -1,- 2).
 Це призводить до багатьох різних варіантів продуктів навіть з таким простим алканом, як гексан. Зауважимо, що циклічні вуглеводні теж піддаються гідроксилювання. У реакції гідроксилювання спочатку утворюється напівацеталь, який потім перетворюється на спирт та альдегід. При окисненні алкенів цитохромом Р450 утворюються двоатомні окиси. Вони відрізняються за своєю стабільністю і можуть бути високо реакційними. Наприклад, вінілхлорид метаболічно переходить в окис, який потім перетворюється на хлорацетальдегід – мутаген, що діє безпосередньо на ДНК. Ці дослідження призвели до заборони використання вінілхлориду в розпилювачах. Вінільна група стерину (вінілбензол) відома своїми канцерогенними властивостями, але організм людини здатний нейтралізувати його, переводячи окис за допомогою ферменту епоксигідролази в діол. Але епоксигідролаза допомагає не завжди. Наприклад, цитохром Р450 синтезує епоксид Афлотоксину В1 in vivо. Ця сполука – високо реакційний електрофіл, не стабільний і швидко формує аддукт з ДНК. До того ж діол, що утворюється з епоксиду, також нестабільний і високо реакційноздатний. Окислення ароматичних сполук цитохромом Р450 також дає епоксиди, але вони швидко переходять у фенол. В результаті гідроксилювання бензолу отриманий фенол може знову гідроксилюватися, переходячи в катехол або гідрохінон. Зауважимо, що катехол та гідрохінон можуть реагувати з киснем, пригнічуючи аналогічні реакції з хінонами та супероксидами, які є токсинами. Така відома сполука як 2,3,7,8-тетрахлордибензолдіоксин (ТХДД) не схильна до гідроксилювання та стійкості (період напіврозпаду в організмі людини – рік і більше).
Це призводить до багатьох різних варіантів продуктів навіть з таким простим алканом, як гексан. Зауважимо, що циклічні вуглеводні теж піддаються гідроксилювання. У реакції гідроксилювання спочатку утворюється напівацеталь, який потім перетворюється на спирт та альдегід. При окисненні алкенів цитохромом Р450 утворюються двоатомні окиси. Вони відрізняються за своєю стабільністю і можуть бути високо реакційними. Наприклад, вінілхлорид метаболічно переходить в окис, який потім перетворюється на хлорацетальдегід – мутаген, що діє безпосередньо на ДНК. Ці дослідження призвели до заборони використання вінілхлориду в розпилювачах. Вінільна група стерину (вінілбензол) відома своїми канцерогенними властивостями, але організм людини здатний нейтралізувати його, переводячи окис за допомогою ферменту епоксигідролази в діол. Але епоксигідролаза допомагає не завжди. Наприклад, цитохром Р450 синтезує епоксид Афлотоксину В1 in vivо. Ця сполука – високо реакційний електрофіл, не стабільний і швидко формує аддукт з ДНК. До того ж діол, що утворюється з епоксиду, також нестабільний і високо реакційноздатний. Окислення ароматичних сполук цитохромом Р450 також дає епоксиди, але вони швидко переходять у фенол. В результаті гідроксилювання бензолу отриманий фенол може знову гідроксилюватися, переходячи в катехол або гідрохінон. Зауважимо, що катехол та гідрохінон можуть реагувати з киснем, пригнічуючи аналогічні реакції з хінонами та супероксидами, які є токсинами. Така відома сполука як 2,3,7,8-тетрахлордибензолдіоксин (ТХДД) не схильна до гідроксилювання та стійкості (період напіврозпаду в організмі людини – рік і більше).



ub-70x70.jpg)