Tipuri de meioză și semnificația ei biologică. Scurtă descriere a etapelor și modelului diviziunii celulare prin meioză Cine a descris pentru prima dată fazele meiozei
Meioza este o metodă specială de împărțire a celulelor eucariote, în care numărul inițial de cromozomi este redus de 2 ori (din grecescul antic „meion” - mai puțin - și de la „meioză” - scădere).
Fazele individuale ale meiozei la animale au fost descrise de V. Flemming (1882), iar la plante de E. Strasburger (1888), iar apoi de savantul rus V.I. Belyaev. Totodată (1887), A. Weissman a fundamentat teoretic necesitatea meiozei ca mecanism de menținere a unui număr constant de cromozomi. Prima descriere detaliată a meiozei în ovocitele de iepure a fost dată de Winyworth (1900).
Deși meioza a fost descoperită cu mai bine de 100 de ani în urmă, studiul meiozei este încă în desfășurare. Interesul pentru meioză a crescut dramatic la sfârșitul anilor 1960, când a devenit clar că aceleași enzime controlate de gene ar putea fi implicate în multe procese legate de ADN. Recent, un număr de biologi au dezvoltat o idee originală: meioza în organismele superioare servește ca garant al stabilității materialului genetic, deoarece în procesul de meioză, atunci când perechile de cromozomi omologi sunt în contact strâns, firele de ADN sunt verificate. este restabilită precizia și deteriorarea care afectează ambele fire. Studiul meiozei a legat metodele și interesele a două științe: citologia și genetica. Acest lucru a condus la nașterea unei noi ramuri de cunoaștere - citogenetica, care este acum strâns legată de biologia moleculară și ingineria genetică.
Semnificația biologică a meiozei constă în următoarele procese:
1. Datorită reducerii numărului de cromozomi ca urmare a meiozei într-un număr de generații în timpul reproducerii sexuale, se asigură constanța numărului de cromozomi.
2. Distribuția independentă a cromozomilor în anafaza primei diviziuni asigură recombinarea genelor aparținând diferitelor grupe de legătură (situate pe diferiți cromozomi). Distribuția meiotică a cromozomilor către celulele fiice se numește segregare cromozomală.
3. Încrucișarea în profaza I a meiozei asigură recombinarea genelor aparținând aceluiași grup de legătură (situat pe același cromozom).
4. Combinația aleatorie a gameților în timpul fertilizării, împreună cu procesele de mai sus, contribuie la variabilitatea genetică.
5. În timpul procesului de meioză are loc un alt fenomen semnificativ. Acesta este procesul de activare a sintezei ARN (sau a activității transcripționale a cromozomilor) în timpul profazei (diploten), asociat cu formarea cromozomilor perie de lampă (se găsesc la animale și unele plante).
Această revenire a profazei la starea de interfază (în timpul mitozei, sinteza ARNm are loc numai în interfază) este o caracteristică specifică a meiozei ca tip special de diviziune celulară.
Trebuie remarcat faptul că protozoarele prezintă o varietate semnificativă de procese meiotice.
În funcție de poziția lor în ciclul de viață, se disting trei tipuri de meioză:
zigotic a-a meioza (inițială) are loc în zigot, adică imediat după fertilizare. Este caracteristic organismelor în al căror ciclu de viață predomină faza haploidă (ascomicete, bisidiomicete, unele alge, sporozoare etc.).
Gametic meioza (terminală) apare în timpul formării gameților. Se observă la animalele pluricelulare (inclusiv la om), precum și la protozoare și unele plante inferioare, în ciclul de viață al cărora predomină faza diploidă.
Intermediar(spori) meioza apare în timpul sporulării la plantele superioare, inclusiv între stadiile de sporofit (plantă) și gametofit (polen, sac embrionar).
Astfel, meioza este o formă de diviziune nucleară însoțită de o scădere a numărului de cromozomi de la diploid la haploid și o modificare a materialului genetic. Rezultatul meiozei este formarea de celule cu un set haploid de cromozomi (celule sexuale).
Durata meiozei poate varia în funcție de tipul de plantă și animal (Tabelul 1).
Tabelul 1. Durata meiozei la diferite specii de plante
Meioza tipică constă din două diviziuni celulare secvenţiale, numite respectiv meioza I şi meioza II. În prima diviziune, numărul de cromozomi scade la jumătate, de aceea se numește prima diviziune meiotică reducţionist, mai rar - heterotipic. În a doua divizie, numărul de cromozomi nu se modifică; această diviziune se numește ecuațională(egalizare), mai rar - homeotipic. Expresiile „meioză” și „diviziune de reducere” sunt adesea folosite în mod interschimbabil.
Numărul inițial de cromozomi din meiocite (celule care intră în meioză) se numește numărul de cromozomi diploid (2n).Numărul de cromozomi din celulele formate ca urmare a meiozei se numește numărul de cromozomi haploid (n). Numărul minim de cromozomi dintr-o celulă se numește numărul de bază (x). Numărul de bază de cromozomi dintr-o celulă corespunde cantității minime de informații genetice (cantitatea minimă de ADN), care se numește genom.
Numărul de genomi dintr-o celulă se numește număr genomic (n). La majoritatea animalelor pluricelulare, la toate gimnospermele și multe angiosperme, conceptul de haploidie-diploidie și conceptul de număr genomic coincid. De exemplu, o persoană are n=x=23 și 2n=2x=46.
Morfologia meiozei - caracteristici ale fazelor
Interfaza
Interfaza pre-meiotică diferă de interfaza normală prin aceea că procesul de replicare a ADN-ului nu ajunge la final: aproximativ 0,2...0,4% din ADN rămâne neduplicat. Astfel, diviziunea celulară începe în stadiul sintetic al ciclului celular. Prin urmare, meioza este numită figurativ mitoză prematură. Cu toate acestea, în general, putem presupune că într-o celulă diploidă (2n) conținutul de ADN este 4c.
În prezența centriolilor, aceștia sunt dublați în așa fel încât celula să aibă doi diplozomi, fiecare dintre care conține o pereche de centrioli.
Prima diviziune meiotică
ADN-ul a suferit replicare. Începe profaza I, cea mai lungă etapă a meiozei.
Faza I este împărțită în următoarele etape:
leptoten - stadiul de filamente subțiri;
zigoten - stadiul de fire duble;
pachiten - stadiul de filamente groase;
diploten - crossing over;
diakineză - dispariția membranei nucleare și a nucleolului.
În profaza timpurie (leptoten), are loc pregătirea pentru conjugarea cromozomilor. Cromozomii au fost deja duplicați, dar cromatidele surori din ei încă nu se pot distinge. Cromozomii încep să se împacheteze.
Spre deosebire de profaza mitozei, în care cromozomii sunt localizați cap la cap de-a lungul membranei nucleare și, atunci când sunt împachetate, sunt atrași de membrană, cromozomii leptotenului cu secțiunile lor telomerice (capete) sunt localizați într-unul dintre polii nucleului, formând un „ buchet” figura la animale și comprimat într-o minge „sineză” - în plante. Această aranjare sau orientare în nucleu permite cromozomilor să realizeze mai rapid și mai ușor conjugarea locilor cromozomiali omologi (Fig. 1).
Evenimentul central este procesul misterios de recunoaștere a cromozomilor omologi și apropierea lor în perechi unul de celălalt are loc în zigotenul profasei I. În timpul conjugării (adunării) cromozomilor omologi, se formează perechi - bivalenți, iar cromozomii sunt scurtați vizibil. Din acest moment începe formarea complexului sinaptonemal (SC). Formarea complexului sinaptonemal și sinopsisul cromozomilor sunt sinonime.
Orez. 1. Etapa profază
În următoarea etapă a profezei I - pahitenăîntre cromozomii omologi există un contact strâns, care se numește sinapsis (de la grecescul synopsis - conexiune, legătură). Cromozomii în acest stadiu sunt foarte spiralați, ceea ce face posibilă observarea lor la microscop.
În timpul sinapsei, omologii se împletesc, adică. conjuga. Bivalenții conjugați sunt legați prin chiasma. Fiecare bivalent este format din doi cromozomi și patru cromatide, unde fiecare cromozom provine de la părintele său. În timpul formării sinapselor (SC), are loc un schimb de regiuni între cromatide omoloage. Acest proces, numit încrucișare, are ca rezultat cromatidele care au acum o compoziție genetică diferită.
Complexul sinaptonemal (SC) atinge cea mai mare dezvoltare în pahiten și în această perioadă este o structură tip panglică situată în spațiul dintre cromozomii omologi paraleli. SC constă din două elemente laterale paralele formate din proteine împachetate dens și un element central mai puțin dens care se extinde între ele (Fig. 2).

Orez. 2. Schema complexului sinaptonemal
Fiecare element lateral este format dintr-o pereche de cromatide surori sub forma axei longitudinale a cromozomului leptoten și, înainte de a deveni parte a SC, se numește element axial. Buclele laterale ale cromatinei se află în afara SC, înconjurând-o pe toate părțile.
Dezvoltarea SC în timpul meiozei:
leptoten - structura cromozomilor care au intrat în leptoten se dovedește imediat a fi neobișnuită: în fiecare omolog există un fir longitudinal care curge de-a lungul axei cromozomilor pe toată lungimea sa;
zigoten - în această etapă, firele axiale ale omologilor se apropie, în timp ce capetele firelor axiale atașate de membrana nucleară par să alunece de-a lungul suprafeței sale interioare unele spre altele;
pahitenă. SC atinge cea mai mare dezvoltare în pachitenă, când toate elementele sale capătă densitate maximă, iar cromatina ia forma unui „înveliș” dens și continuu în jurul său.
Funcții SC:
1. Un complex sinaptonemal complet dezvoltat este necesar pentru reținerea normală a omologilor în bivalent atât timp cât este necesar pentru încrucișarea și stabilirea chiasmelor. Cromozomii sunt conectați folosind complexul sinaptonemal pentru o perioadă de timp (de la 2 ore în drojdie până la 2-3 zile la om), timp în care se fac schimb de secțiuni de ADN omolog între cromozomi omologi - crossing over (din engleză, crossing over - formarea unei încrucișări) .
2. Prevenirea conexiunii prea puternice a omologilor și menținerea lor la o anumită distanță, păstrarea individualității lor, creând oportunitatea de a împinge în diploten și de a se dispersa în anafază.
Procesul de încrucișare este asociat cu activitatea anumitor enzime, care, atunci când se formează chiasma între cromatidele surori, le „taie” în punctul de încrucișare, urmată de reunificarea fragmentelor rezultate. În cele mai multe cazuri, aceste procese nu duc la nicio perturbare în structura genetică a cromozomilor omologi, de exemplu. are loc conexiunea corectă a fragmentelor de cromatide și se reface structura lor originală.
Cu toate acestea, este posibilă și o altă versiune (rară) a evenimentelor, care este asociată cu reunificarea eronată a fragmentelor de structuri tăiate. În acest caz, are loc un schimb reciproc de secțiuni de material genetic între cromatidele de conjugare (recombinare genetică).
În fig. Figura 3 prezintă o diagramă simplificată a unor variante posibile de încrucișare simplă sau dublă care implică două cromatide dintr-o pereche de cromozomi omologi. Trebuie subliniat că încrucișarea este un eveniment aleatoriu care, cu probabilitate variabilă, poate apărea pe orice secțiune (sau pe două sau mai multe secțiuni) de cromozomi omologi. În consecință, în stadiul de maturare a gameților unui organism eucariot în profaza primei diviziuni a meiozei, funcționează principiul universal al combinării aleatorii (libere) (recombinării) a materialului genetic al cromozomilor omologi.
În studiile citologice ale sinapselor din ultimele două decenii, un rol important l-a jucat metoda de răspândire a celulelor meiotice profază de animale și plante sub influența unei soluții hipotonice. Metoda a intrat în citogenetică după munca lui Moise și a jucat același rol ca și metoda de pregătire a preparatelor „presate” pentru studiul cromozomilor metafazici jucată odată, salvând citogeneticienii de secțiunile de microtom.
Metoda Moses și modificările sale au devenit mai convenabile decât analiza SC pe secțiuni ultrasubțiri. Această metodă a stat la baza studiilor asupra meiozei și a acoperit treptat problemele controlului genetic al meiozei la animale și plante.

Orez. 3. Variante separate de încrucișare simplă și dublă care implică două cromatide: 1 cromatidă originală și o variantă fără încrucișare; 2 cromatide de încrucișare unică în zona A B și cromatide de încrucișare; 3 cromatide de încrucișare unică în regiunea B-C și cromatide de încrucișare; 4 cromatide duble încrucișate și încrucișate în mai multe zone diferite, bazate pe omologia materialului genetic al acestor zone. Se crede că pe fiecare parte, fie una dintre cele două cromatide surori ale cromozomului corespunzător, fie ambele cromatide pot participa la procesul de conjugare.
În dipotenă, cromozomii omologi, după împerechere și încrucișare, încep să se respingă unul pe altul. Procesul de repulsie începe la centromer. Divergența omologilor este împiedicată de chiasmata - joncțiunea cromatidelor non-surori rezultată din încrucișare. Pe măsură ce cromatidele diverg, unele chiasmate se deplasează spre capătul brațului cromozomului. De obicei, există mai multe încrucișări și cu cât cromozomii sunt mai lungi, cu atât mai mulți dintre ei, prin urmare, în diploten, de regulă, există mai multe chiasmate într-un singur bivalent.
În timpul etapei de diakineză, numărul chiasmelor scade. Bivalenții sunt localizați de-a lungul periferiei nucleului. Nucleolul se dizolvă, membrana este distrusă și începe trecerea la metafaza I. Pe tot parcursul profezei se păstrează nucleolul și învelișul nuclear. Înainte de profază, în perioada sintetică a interfazei, au loc replicarea ADN-ului și reproducerea cromozomilor. Cu toate acestea, această sinteză nu se încheie complet: ADN-ul este sintetizat cu 99,8%, iar proteinele cu 75%. Sinteza ADN-ului se termină în pahitenă, sinteza proteinelor în diploten.
În metafaza I, structura în formă de fus formată din microtubuli devine vizibilă. În timpul meiozei, microtubulii individuali sunt atașați de centromerii cromozomilor fiecărui bivalent. Apoi perechile de cromozomi se deplasează în planul ecuatorial al celulei, unde sunt aranjați într-o ordine aleatorie. Centromerii cromozomilor omologi sunt situati pe laturile opuse ale planului ecuatorial; în metafaza mitozei, dimpotrivă, centromerii cromozomilor individuali sunt localizați în planul ecuatorial.
În metafaza I, bivalenții sunt localizați în centrul celulei, în zona plăcii ecuatoriale (Fig. 4).

Orez. 4. Stadiile meiozei: profaza I - metafaza I
Anafaza începe cu divergența cromozomilor omologi și mișcarea lor spre poli. În cromozomii fără centromer, atașarea nu poate exista. În anafaza mitozei, centromerii se divid și cromatide identice se separă. În anafaza I a meiozei, centromerii nu se divid, cromatidele rămân împreună, iar cromozomii omologi sunt separați. Cu toate acestea, din cauza schimbului de fragmente ca urmare a încrucișării, cromatidele nu sunt identice, ca la începutul meiozei. În anafaza I, omologii conjugați se deplasează spre poli.
În celulele fiice, numărul de cromozomi este la jumătate mai mare (mult haploid), în timp ce masa ADN-ului este, de asemenea, redusă la jumătate, iar cromozomii rămân dicromatizi. Divergența precisă a perechilor omoloage la poli opuși stă la baza reducerii numărului lor.
În telofaza I, cromozomii se concentrează la poli și suferă o oarecare decondensare, din cauza căreia spiralarea cromozomilor slăbește, aceștia se alungesc și din nou devin nedistinși (Fig. 5). Pe măsură ce telofaza trece treptat în interfază, învelișul nuclear (inclusiv fragmente ale membranei nucleare a celulei mamă), precum și septul celular, iese din reticulul endoplasmatic. În cele din urmă, se formează din nou nucleolul și se reia sinteza proteinelor.

Orez. 5. Stadiile meiozei: anafaza I - telofaza I
În interkineză, se formează nuclei, fiecare dintre care conține n cromozomi dicromatidici.
Particularitatea celei de-a doua diviziuni a meiozei este, în primul rând, că în interfaza II nu are loc dublarea cromatinei, prin urmare fiecare celulă care intră în profaza II păstrează același raport n2c.
A doua diviziune meiotică
În timpul celei de-a doua diviziuni a meiozei, cromatidele surori ale fiecărui cromozom se deplasează spre poli. Deoarece încrucișarea ar putea avea loc în profaza I, iar cromatidele surori ar putea deveni neidentice, se obișnuiește să spunem că a doua diviziune are loc ca mitoză, dar aceasta nu este o mitoză adevărată, în care celulele fiice conțin în mod normal cromozomi identici ca formă și set de gene.
La începutul celei de-a doua diviziuni meiotice, cromatidele sunt încă conectate prin centromeri. Această diviziune este similară mitozei: dacă s-a format o membrană nucleară în telofaza I, acum este distrusă, iar la sfârșitul scurtei profase II nucleolul dispare.

Orez. 6. Stadiile meiozei: profaza II-metafaza II
În metafaza II, fusul și cromozomii, formați din două cromatide, pot fi văzute din nou. Cromozomii sunt atașați prin centromeri de firele fusului și se aliniază în planul ecuatorial (Fig. 6). În anafaza II, centromerii se divid și se separă, iar cromatidele surori, acum cromozomi, se deplasează la poli opuși. În telofaza II se formează noi membrane nucleare și nucleoli, comprimarea cromozomilor slăbește și aceștia devin invizibili în nucleul interfazic (Fig. 7).

Orez. 7. Stadiile meiozei: anafaza II - telofaza II
Meioza se termină cu formarea de celule haploide - gameți, tetrade de spori - descendenți ai celulei originale cu un set de cromozomi (haploid) înjumătățit și o masă de ADN haploid (celula originală 2n, 4c, - spori, gameți - n, c) .
Diagrama generală a distribuției cromozomilor unei perechi omoloage și a celor două perechi de gene alelice diferite pe care le conțin în timpul a două diviziuni meiotice este prezentată în Fig. 8. După cum se poate vedea din această diagramă, sunt posibile două opțiuni fundamental diferite pentru o astfel de distribuție. Prima variantă (mai probabilă) este asociată cu formarea a două tipuri de gameți diferiți genetic cu cromozomi care nu au suferit încrucișări în zonele în care sunt localizate genele în cauză. Astfel de gameți sunt de obicei numiți non-crossover. În a doua opțiune (mai puțin probabilă), împreună cu gameții neîncrucișați, gameții încrucișați apar și ca urmare a schimbului genetic (recombinare genetică) în secțiuni de cromozomi omologi situate între loci a două gene non-alelice.

Orez. 8. Două variante ale distribuției cromozomilor unei perechi omoloage și genele non-alelice pe care le conțin ca urmare a două diviziuni meiotice
Meioză
Concepte de bază și definiții
Meioza este o metodă specială de împărțire a celulelor eucariote, în care numărul inițial de cromozomi este redus de 2 ori (din greaca veche „ meyon" - mai puțin - și de la " meioză" - scădea). Deseori se numește o scădere a numărului de cromozomi reducere.
Numărul inițial de cromozomi în meiocite(celule care intră în meioză) se numește numărul de cromozomi diploid (2n) Numărul de cromozomi din celulele formate ca urmare a meiozei se numește numărul cromozomilor haploid (n).
Numărul minim de cromozomi dintr-o celulă se numește numărul de bază ( X). Numărul de bază de cromozomi dintr-o celulă corespunde cantității minime de informații genetice (cantitatea minimă de ADN), care se numește genă O m. Numărul de gene O mov într-o celulă se numește genă O multe numere (Ω). La majoritatea animalelor pluricelulare, la toate gimnospermele și multe angiosperme, conceptul de haploidie-diploidie și conceptul de genă O multe numere se potrivesc. De exemplu, la o persoană n=X=23 și 2 n=2X=46.
Caracteristica principală a meiozei este conjugare(pereche) cromozomi omologi cu divergenţa lor ulterioară în celule diferite. Distribuția meiotică a cromozomilor către celulele fiice se numește segregarea cromozomilor.
O scurtă istorie a descoperirii meiozei
Fazele individuale ale meiozei la animale au fost descrise de V. Flemming (1882), iar la plante de E. Strasburger (1888), iar apoi de savantul rus V.I. Belyaev. Totodată (1887), A. Weissman a fundamentat teoretic necesitatea meiozei ca mecanism de menținere a unui număr constant de cromozomi. Prima descriere detaliată a meiozei în ovocitele de iepure a fost dată de Winyworth (1900). Studiul meiozei este încă în curs de desfășurare.
Cursul general al meiozei
Meioza tipică constă din două diviziuni celulare succesive, care se numesc respectiv meioza IȘi meioza II. În prima diviziune, numărul de cromozomi scade la jumătate, de aceea se numește prima diviziune meiotică reducţionist, mai rar - heterotipic. În a doua divizie, numărul de cromozomi nu se modifică; această diviziune se numește ecuațională(egalizare), mai rar - homeotipic. Expresiile „meioză” și „diviziune de reducere” sunt adesea folosite în mod interschimbabil.
Interfaza
Interfaza pre-meiotică diferă de interfaza obișnuită prin aceea că procesul de replicare a ADN-ului nu ajunge la final: aproximativ 0,2...0,4% din ADN rămâne neduplicat. Astfel, diviziunea celulară începe în stadiul sintetic al ciclului celular. Prin urmare, meioza este numită figurativ mitoză prematură. Cu toate acestea, în general, putem presupune că într-o celulă diploidă (2 n) Conținutul de ADN este 4 Cu.
În prezența centriolilor, aceștia sunt dublați în așa fel încât celula să aibă doi diplozomi, fiecare dintre care conține o pereche de centrioli.
Prima diviziune a meiozei (diviziune de reducere, sau meioza I)
Esența diviziunii de reducere este reducerea la jumătate a numărului de cromozomi: din celula diploidă originală se formează două celule haploide cu cromozomi bicromatidici (fiecare cromozom include 2 cromatide).
Profaza 1(profaza primei divizii) constă dintr-un număr de etape:
Leptoten(etapa de fire subtiri). Cromozomii sunt vizibili la microscopul luminos sub forma unei mingi de fire subtiri. Se numește leptoten timpuriu, când firele de cromozomi sunt încă foarte puțin vizibile proleptoten.
Zigoten(etapa de îmbinare a firelor). Se întâmplă conjugarea cromozomilor omologi(din lat. conjugare– conectare, împerechere, fuziune temporară). Cromozomii omologi (sau omologii) sunt cromozomi care sunt similari între ei din punct de vedere morfologic și genetic. În organismele diploide normale, cromozomii omologi sunt perechi: organismul diploid primește un cromozom de la pereche de la mamă, iar celălalt de la tată. În timpul conjugării, ele se formează bivalente. Fiecare bivalent este un complex relativ stabil de o pereche de cromozomi omologi. Omologii sunt ținuți împreună de proteinele proteice. complexe sinaptonemale. Un complex sinaptonemal poate conecta doar două cromatide la un punct. Numărul de bivalenți este egal cu numărul haploid de cromozomi. În caz contrar, se numesc bivalenți tetrade, deoarece fiecare bivalent include 4 cromatide.
Pachytena(etapa cu filament gros). Cromozomii sunt spiralați, iar eterogenitatea lor longitudinală este clar vizibilă. Replicarea ADN-ului este finalizată (o specială ADN pahiten). Se termină trecere peste- încrucișarea cromozomilor, în urma căreia fac schimb de secțiuni de cromatide.
Diplotena(etapă cu filet dublu). Cromozomii omologi din bivalenți se resping reciproc. Ele sunt conectate în puncte separate numite chiasmata(din litera greacă veche χ - „chi”).
Diacineza(stadiul divergenței bivalente). Bivalenții individuali sunt localizați la periferia nucleului.
Metafaza I(metafaza prima divizie)
ÎN prometafaza I membrana nucleară este distrusă (fragmentată). Se formează fusul de fisiune. În continuare, apare metakineza - bivalenții se deplasează în planul ecuatorial al celulei.
Anafaza I(anafaza primei divizii)
Cromozomii omologi care alcătuiesc fiecare bivalent sunt separați, iar fiecare cromozom se deplasează spre cel mai apropiat pol al celulei. Separarea cromozomilor în cromatide nu are loc. Procesul de distribuire a cromozomilor la celulele fiice se numește segregarea cromozomilor.
Telofaza I(telofaza primei divizii)
Cromozomii bicromatidici omologi diverg complet către polii celulari. În mod normal, fiecare celulă fiică primește un cromozom omolog de la fiecare pereche de omologi. Se formează două haploid nuclee care conțin jumătate de cromozomi decât nucleul celulei diploide originale. Fiecare nucleu haploid conține un singur set de cromozomi, adică fiecare cromozom este reprezentat de un singur omolog. Conținutul de ADN din celulele fiice este 2 Cu.
În majoritatea cazurilor (dar nu întotdeauna), telofaza I este însoțită de citokineza .
Interkineza
Interkineza este intervalul scurt dintre două diviziuni meiotice. Diferă de interfază prin aceea că nu au loc replicarea ADN-ului, duplicarea cromozomilor și duplicarea centriolului: aceste procese au avut loc în interfaza pre-meiotică și, parțial, în profaza I.
A doua diviziune a meiozei (diviziunea ecuației, sau meioza II)
În timpul celei de-a doua diviziuni a meiozei, numărul de cromozomi nu scade. Esența diviziunii ecuaționale este formarea a patru celule haploide cu cromozomi cu o singură cromatidă (fiecare cromozom conține o cromatidă).
Profaza II(profaza diviziei a doua)
Nu diferă semnificativ de profaza mitozei. Cromozomii sunt vizibili la microscopul luminos sub formă de fire subțiri. În fiecare dintre celulele fiice se formează un fus de diviziune.
Metafaza II(metafaza diviziei a doua)
Cromozomii sunt localizați în planurile ecuatoriale ale celulelor haploide, independent unul de celălalt. Aceste planuri ecuatoriale pot fi situate în același plan, pot fi paralele între ele sau reciproc perpendiculare.
Anafaza II(anafaza a doua divizie)
Cromozomii sunt separați în cromatide (ca și în mitoză). Cromozomii cu o singură cromatidă rezultați, ca parte a grupurilor anafazate, se deplasează la polii celulelor.
Telofaza II(telofaza a doua divizie)
Cromozomii cu o singură cromatidă s-au mutat complet la polii celulei și se formează nuclei. Conținutul de ADN din fiecare celulă devine minim și se ridică la 1 Cu.
Tipuri de meioză și semnificația ei biologică
În general, meioza produce patru celule haploide dintr-o celulă diploidă. La meioză gametică gameții se formează din celulele haploide rezultate. Acest tip de meioză este caracteristic animalelor. Meioza gametică este strâns legată de gametogenezaȘi fertilizare. La zigoticȘi meioza sporilor celulele haploide rezultate dau naştere la spori sau zoospori. Aceste tipuri de meioză sunt caracteristice eucariotelor inferioare, ciupercilor și plantelor. Meioza sporilor este strâns legată de sporogeneza. Prin urmare, meioza este baza citologică a reproducerii sexuale și asexuate (spori)..
Semnificația biologică a meiozei este de a menține un număr constant de cromozomi în prezența procesului sexual. În plus, din cauza traversării, recombinare– apariţia unor noi combinaţii de înclinaţii ereditare în cromozomi. Meioza oferă, de asemenea variabilitate combinativă– apariția de noi combinații de înclinații ereditare în timpul fertilizării ulterioare.
Cursul meiozei este controlat de genotipul organismului, sub controlul hormonilor sexuali (la animale), al fitohormonilor (la plante) și al multor alți factori (de exemplu, temperatura).
Meioză (din greaca meioză- reducerea) este un tip special de diviziune a celulelor eucariote, în care, după o singură dublare a ADN-ului, celula împărțit de două ori , iar dintr-o celulă diploidă se formează 4 haploide. Constă din 2 divizii consecutive (desemnate I și II); fiecare dintre ele, ca si mitoza, include 4 faze (profaza, metafaza, anafaza, telofaza) si citokineza.
Fazele meiozei:
Profaza eu , este complex, împărțit în 5 etape:
1. leptonem (din greaca leptos- subțire, nema– fir) – cromozomii se spiralează și devin vizibili ca fire subțiri. Fiecare cromozom omolog a fost deja replicat cu 99,9% și constă din două cromatide surori legate între ele la centromer. Conținutul materialului genetic - 2 n 2 xp 4 c. Cromozomii cu ajutorul grupurilor de proteine ( discuri de atașare ) sunt atașate la ambele capete de membrana interioară a învelișului nuclear. Învelișul nuclear este păstrat, nucleolul este vizibil.
2. Zygonema (din greaca zigon – pereche) – cromozomii diploizi omologi se repezi unul spre celălalt și se conectează mai întâi în regiunea centromerului, apoi pe toată lungimea ( conjugare ). Sunt formate bivalente (din lat. bi – dublu, valens– puternic), sau tetrade cromatidă. Numărul de bivalenți corespunde setului haploid de cromozomi; conținutul materialului genetic poate fi scris ca 1 n 4 xp 8 c. Fiecare cromozom dintr-un bivalent provine fie de la tată, fie de la mamă. Cromozomi sexuali situat în apropierea membranei nucleare interioare. Această zonă se numește veziculă genitală.
Între cromozomi omologi în fiecare bivalent, specializat complexe sinaptonemale (din greaca sinapsa– legătură, legătură), care sunt structuri proteice. La o mărire mare, două fire paralele de proteine, fiecare cu grosimea de 10 nm, sunt vizibile în complex, conectate prin dungi subțiri transversale de aproximativ 7 nm în dimensiune, pe ambele părți ale cărora se află cromozomi sub formă de multe bucle.
In centrul complexului se afla element axial grosime 20 – 40 nm. Complexul sinaptonemal este comparat cu scara de franghie , ale căror laturi sunt formate din cromozomi omologi. O comparație mai exactă - Fermoar .
Până la sfârșitul zigonemului, fiecare pereche de cromozomi omologi este conectată între ele folosind complexe sinaptonemale. Numai cromozomii sexuali X și Y nu se conjugă complet, deoarece nu sunt complet omologi.
3. B pachyneme (din greaca pahys– gros) bivalenții se scurtează și se îngroașă. Între cromatidele de origine maternă și paternă, conexiunile apar în mai multe locuri - chiasmata (din greacă.c hiazma- cruce). În zona fiecărei chiasme, un complex de proteine implicate în recombinare (d~ 90 nm), iar schimbul de secțiuni corespunzătoare de cromozomi omologi are loc - de la patern la matern și invers. Acest proces se numește crossover (din engleza Curosing- peste– răscruce de drumuri). În fiecare om bivalent, de exemplu, trecerea are loc în două până la trei zone.
4. B diplomă (din greaca diploos– complexele sinaptonemale duble se dezintegrează, iar cromozomii omologi ai fiecărui bivalent îndepărtați unul de celălalt, dar legătura dintre ele rămâne în zonele chiasmatei.
5. Diacineza (din greaca diakineina- A trece prin). În diakineză, condensarea cromozomilor este completă, aceștia sunt separați de membrana nucleară, dar cromozomii omologi continuă să rămână legați între ei prin secțiuni terminale, iar cromatidele surori ale fiecărui cromozom prin centromeri. Bivalenții capătă o formă bizară inele, cruci, opturi etc. În acest moment, membrana nucleară și nucleolii sunt distruse. Centriolii replicați sunt direcționați către poli, iar firele fusului sunt atașate de centromerii cromozomilor.
În general, profaza meiotică este foarte lungă. Când se dezvoltă sperma, poate dura câteva zile, iar când se dezvoltă ovule, poate dura mulți ani.
Metafaza eu seamănă cu un stadiu similar al mitozei. Cromozomii sunt instalați în planul ecuatorial, formând placa metafazică. Spre deosebire de mitoză, microtubulii fusului sunt atașați de centromerul fiecărui cromozom doar pe o parte (partea polului), iar centromerii cromozomilor omologi sunt localizați pe ambele părți ale ecuatorului. Legătura dintre cromozomi cu ajutorul chiasmei continuă să fie păstrată.
ÎN anafaza eu chiasma se dezintegrează, cromozomii omologi se separă unul de celălalt și diverg către poli. Centromeri dintre acești cromozomi, însă, spre deosebire de anafaza mitozei, nu sunt replicate, ceea ce înseamnă că cromatidele surori nu se separă. Divergenta cromozomilor este natura aleatorie. Conţinutul informaţiei genetice devine 1 n 2 xp 4 c la fiecare pol al celulei și în celula ca întreg - 2(1 n 2 xp 4 c) .
ÎN telofaza eu , ca și în mitoză, se formează, se formează și se adâncesc membranele nucleare și nucleolii brazdă de clivaj. Apoi se întâmplă citokineza . Spre deosebire de mitoză, derularea cromozomilor nu are loc.
Ca urmare a meiozei I, se formează 2 celule fiice care conțin un set haploid de cromozomi; fiecare cromozom are 2 cromatide (recombinante) distincte genetic: 1 n 2 xp 4 c. Prin urmare, ca urmare a meiozei apare I reducere (reducerea la jumătate) a numărului de cromozomi, de unde și numele primei diviziuni - reducere .
După sfârșitul meiozei I există o perioadă scurtă - interkineza , timp în care nu au loc replicarea ADN-ului și duplicarea cromatidelor.
Profaza II nu durează mult, iar conjugarea cromozomilor nu are loc.
ÎN metafaza II cromozomii se aliniază în planul ecuatorial.
ÎN anafaza II ADN-ul din regiunea centromerului este replicat, așa cum se întâmplă în anafaza mitozei, cromatidele se deplasează spre poli.
După telofaze II Și citokineza II celulele fiice se formează conținând material genetic în fiecare - 1 n 1 xp 2 c. În general, se numește a doua divizie ecuațională (egalizarea).
Deci, în urma a două diviziuni meiotice succesive, se formează 4 celule, fiecare poartă un set haploid de cromozomi.
Meioza apare în celulele organismelor care se reproduc sexual.
Sensul biologic al fenomenului este determinat de un nou set de caracteristici la descendenți.
În această lucrare, vom lua în considerare esența acestui proces și, pentru claritate, o vom prezenta în figură, ne vom uita la succesiunea și durata diviziunii celulelor germinale și vom afla, de asemenea, asemănările și diferențele dintre mitoza si meioza.
Ce este meioza
Un proces însoțit de formarea a patru celule cu un singur cromozom stabilit dintr-unul original.

Informația genetică a fiecărei celule nou formate corespunde cu jumătate din setul de celule somatice.
Fazele meiozei
Diviziunea meiotică implică două etape, formate fiecare din patru faze.
Prima divizie
Include profaza I, metafaza I, anafaza I și telofaza I.

Profaza I
În această etapă, se formează două celule cu jumătate din setul de informații genetice. Profaza primei divizii include mai multe etape. Este precedat de interfaza pre-meiotică, în timpul căreia are loc replicarea ADN-ului.
Apoi are loc condensarea, formarea de filamente lungi și subțiri cu o axă proteică în timpul leptotenului. Acest fir este atașat de membrana nucleară cu ajutorul unor extensii terminale - discuri de atașare. Jumătățile cromozomilor duplicați (cromatide) nu sunt încă distinse. Când sunt examinate, arată ca niște structuri monolitice.
Urmează etapa zigotenului. Omologii fuzionează pentru a forma bivalenți, al căror număr corespunde unui singur număr de cromozomi. Procesul de conjugare (conectare) se realizează între perechi asemănătoare din punct de vedere genetic și morfologic. Mai mult, interacțiunea începe de la capete, răspândindu-se de-a lungul corpurilor cromozomiale. Un complex de omologi legați de o componentă proteică - o bivalentă sau tetradă.
Spiralizarea are loc în timpul etapei de filament gros, pachiten. Aici duplicarea ADN-ului a fost deja complet complet și începe trecerea. Acesta este un schimb de regiuni omoloage. Ca rezultat, se formează gene legate cu informații genetice noi. Transcrierea are loc în paralel. Secțiuni dense de ADN - cromomeri - sunt activate, ceea ce duce la o modificare a structurii cromozomilor, cum ar fi „periile lămpii”.
Cromozomii omologi se condensează, se scurtează și diverg (cu excepția punctelor de legătură - chiasma). Aceasta este o etapă în biologia diplotenului sau dictiotenului. Cromozomii în acest stadiu sunt bogați în ARN, care este sintetizat în aceleași zone. În ceea ce privește proprietățile, acesta din urmă este aproape de informațional.
În cele din urmă, bivalenții se dispersează la periferia nucleului. Acestea din urmă se scurtează, își pierd nucleolii și devin compacte, neasociate cu învelișul nuclear. Acest proces se numește diakineză (tranziția la diviziunea celulară).
Metafaza I
Apoi, bivalenții se deplasează spre axa centrală a celulei. Fusele de diviziune se extind de la fiecare centromer, fiecare centromer este echidistant de ambii poli. Mișcări de amplitudine mică ale firelor le țin în această poziție.
Anafaza I
Cromozomii, formați din două cromatide, se separă. Recombinarea are loc cu scăderea diversității genetice (datorită absenței omologilor în setul de gene situate în loci (regiuni).
Telofaza I
Esența fazei este divergența cromatidelor cu centromerii lor către părți opuse ale celulei. Într-o celulă animală are loc diviziunea citoplasmatică, într-o celulă vegetală are loc formarea unui perete celular.
Divizia a doua
După interfaza primei diviziuni, celula este pregătită pentru a doua etapă.

Profaza II
Cu cât telofaza este mai lungă, cu atât durata profasei este mai scurtă. Cromatidele se aliniază de-a lungul celulei, formând un unghi drept cu axele lor în raport cu firele primei diviziuni meiotice. În această etapă, se scurtează și se îngroașă, iar nucleolii suferă dezintegrare.
Metafaza II
Centromerii sunt din nou localizați în planul ecuatorial.
Anafaza II
Cromatidele se separă unele de altele, îndreptându-se spre poli. Acum se numesc cromozomi.
Telofaza II
Despiralizarea, întinderea cromozomilor formați, dispariția fusului, dublarea centriolilor. Nucleul haploid este înconjurat de o membrană nucleară. Se formează patru celule noi.
Tabel de comparație între mitoză și meioză
Caracteristicile și diferențele sunt prezentate pe scurt și clar în tabel.
| Caracteristici | Diviziune meiotică | Diviziunea mitotică |
| Numărul de diviziuni | realizat în două etape | realizat într-o singură etapă |
| Metafaza | după duplicare, cromozomii sunt aranjați în perechi de-a lungul axei centrale a celulei | după duplicare, cromozomii sunt localizați individual de-a lungul axei centrale a celulei |
| Fuziune | Există | Nu |
| Trecere peste | Există | Nu |
| Interfaza | fără duplicare ADN în interfaza II | Dublarea ADN-ului are loc înainte de divizare |
| Rezultatul diviziunii | gameti | somatic |
| Localizare | în gameţi în curs de maturizare | în celulele somatice |
| Calea de redare | sexual | asexuat |
Datele prezentate sunt o diagramă a diferențelor, iar asemănările se reduc la aceleași faze, reduplicarea ADN-ului și helicoidalizarea înainte de începerea ciclului celular.
Semnificația biologică a meiozei

Care este rolul meiozei:
- Oferă noi combinații de gene datorită încrucișării.
- Sprijină variabilitatea combinativă. Meioza este o sursă de noi trăsături într-o populație.
- Menține un număr constant de cromozomi.
Concluzie
Meioza este un proces biologic complex în timpul căruia se formează patru celule, cu noi caracteristici obținute ca urmare a încrucișării.
Nikolay Mushkambarov, Dr. biol. stiinte
Omenirea îmbătrânește, dar toată lumea vrea să trăiască nu doar mult timp, ci și fără acele boli care vin odată cu vârsta. În ultima jumătate de secol, au apărut multe teorii „revoluționare” ale îmbătrânirii, aproape toate oferind o modalitate sigură și fiabilă de a încetini sau chiar de a opri timpul. În fiecare an apar noi senzații, noi descoperiri și noi declarații, încurajatoare și promițătoare. Bioregulatori peptidici, elixir de longevitate, ioni dătătoare de viață sau antioxidant SkQ. Fugi la farmacie, plătește și trăiește, conform instrucțiunilor incluse, până la 100-120 de ani! În ce măsură poți avea încredere în descoperiri senzaționale și care este „adevărul despre îmbătrânire”?
Profesorul N. N. Mushkambarov. Fotografie de Andrey Afanasyev.
August Weismann (1834-1914) - zoolog și evoluționist german. A creat o teorie conform căreia caracteristicile ereditare sunt păstrate și transmise prin germoplasmă fără vârstă.
Leonard Hayflick este un microbiolog american. În anii 1960, el a descoperit că în condiții de laborator, celulele umane și animale se pot diviza doar de un număr limitat de ori.
Alexey Matveevich Olovnikov este un biochimist rus. Pentru a explica experimentele lui Hayflick din 1971, el a prezentat o ipoteză despre scurtarea secțiunilor terminale ale cromozomilor (telomerilor) cu fiecare diviziune celulară.
Știință și viață // Ilustrații
Elizabeth Blackburn și Carol Greider sunt biologi americani. În 1985, a fost descoperită enzima telomeraza. Mecanismul de acțiune al telomerazei este codificarea repetată a noilor secvențe de nucleotide la secțiunile terminale ale telomerilor și restaurarea lor originală.
Benjamin Gompertz (1779-1865) - matematician britanic. El a propus o funcție care descrie statisticile mortalității umane în funcție de vârstă. Această funcție a fost utilizată pentru evaluarea riscurilor în asigurările de viață.
Cartea lui M. M. Vilenchik „Bazele biologice ale îmbătrânirii și longevității”, publicată în 1976, a fost una dintre primele cărți de știință populare pe tema îmbătrânirii și s-a bucurat de un succes enorm.
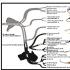
Schema meiozei (folosind exemplul unei perechi de cromozomi omologi). În profaza primei diviziuni a meiozei, cromozomii sunt dublați; apoi cromozomii omologi se conjugă între ei și, menținându-și activitatea, intră în crossover.
Doctor în științe biologice, profesor al Departamentului de Histologie de la Universitatea Medicală de Stat din Moscova, numit după N.V. I. M. Secenov Nikolay Mushkambarov.
Nikolai Nikolaevici, critici aspru multe prevederi binecunoscute ale gerontologiei moderne. Vă rugăm să subliniați obiectele criticii dvs.
Sunt mai mult decât suficiente obiecte! De exemplu, acum este la modă să ne referim la Weisman aproape ca fiind adevărul suprem. Acesta este un biolog celebru care, în secolul al XIX-lea, a postulat că îmbătrânirea nu a apărut imediat în evoluție, ci doar la un moment dat ca un fenomen adaptativ. Din aceasta au ajuns la concluzia că trebuie să existe specii care nu îmbătrânesc: în primul rând, cele mai primitive organisme. În același timp, ei uită cumva că dacă nu îmbătrânesc, atunci trebuie să aibă reparație 100% ADN-ului. Acesta este printre cele mai primitive! Cumva unul nu se potrivește cu celălalt.
Există un mit asociat cu numele unui alt biolog celebru - Leonard Hayflick. Din anii șaizeci ai secolului trecut, lumea științifică a fost încrezătoare că celulele somatice umane au o limită de 50 de diviziuni, iar o astfel de limită în biologie se numește „limita Hayflick”. Cu aproximativ douăzeci de ani în urmă, au fost izolate celule stem care se presupune că erau capabile de un număr nelimitat de diviziuni. Și acest mit (50 pentru toată lumea și infinit pentru celulele stem) persistă în minte până astăzi. De fapt, celulele stem, după cum se dovedește, îmbătrânesc (adică infinitul este abolit) și nu este deloc clar unde să numărăm aceste 50 de diviziuni. Este atât de neclar încât, cel mai probabil, nu există o singură limită de diviziune care să fie universală pentru toate celulele umane în diviziune.
- Ei bine, cum rămâne cu teoria telomerilor a îmbătrânirii? Te face și ea să fii neîncrezător?
Acesta este cel mai popular mit. Conform acestei teorii, întregul mecanism al îmbătrânirii se reduce la faptul că celulele care se divizează sunt lipsite de enzima telomeraza, care prelungește capetele cromozomilor (aceste capete se numesc telomeri) și, prin urmare, cu fiecare diviziune, telomerii sunt scurtați cu 50- 100 de perechi de nucleotide ADN. Enzima telomeraza există, iar descoperirea ei a fost distinsă cu Premiul Nobel în 2009. Iar fenomenul de scurtare a cromozomilor în celulele în diviziune lipsite de telomerază este, de asemenea, fără îndoială (deși se datorează unui motiv ușor diferit de cel subliniat de autorul teoriei telomerilor, Alexey Olovnikov). Dar a reduce îmbătrânirea la acest fenomen este același lucru cu înlocuirea celei mai complexe partituri simfonice cu note de bătaie pe o tobă. Nu întâmplător în 2003 A. Olovnikov și-a abandonat public teoria, înlocuind-o cu așa-zisa teorie reumeric (de asemenea, de altfel, indiscutabilă). Dar și astăzi, chiar și în universitățile de medicină, cursurile de biologie prezintă teoria telomerilor drept cea mai recentă realizare a gândirii științifice. Acest lucru este, desigur, absurd.
Un alt exemplu vine din statisticile mortalității. Formula principală pentru această statistică este ecuația Gompertz, propusă în 1825, sau, cu un termen de corecție, ecuația Gompertz-Makem (1860). Aceste ecuații au doi și, respectiv, trei coeficienți, iar valorile coeficienților variază foarte mult între diferitele populații de oameni. Și se dovedește că modificările coeficienților fiecărei ecuații se corelează între ele. Pe baza cărora se formulează modele globale, la nivel mondial: așa-numita corelație Strehler-Mildvan și efectul compensator al mortalității care a înlocuit-o în acest post - ipoteza soților Gavrilov.
Am compilat un mic model pentru o populație condiționată de oameni și cu ajutorul lui m-am convins că toate aceste modele sunt cel mai probabil un artefact. Faptul este că o mică eroare în determinarea unui coeficient creează o abatere bruscă de la valoarea reală a altui coeficient. Și aceasta este percepută (în coordonate semi-logaritmice) ca o corelație semnificativă din punct de vedere biologic și servește ca o promisiune pentru concluzii bine gândite.
- Ești sigur că ai dreptate când vorbești despre artefact?
Desigur că nu! În general, este dăunător pentru oamenii de știință să fie absolut siguri de ceva, deși există o mulțime de astfel de exemple. Dar am făcut tot posibilul să verific contrariul: că corelațiile nu sunt un artefact. Și nu am putut verifica acest lucru opus. Deci, deocamdată, pe baza unei analize personale, foarte modeste ca amploare, am mai multe motive să cred că corelațiile numite sunt încă artificiale. Ele reflectă erorile metodei și nu modelele biologice.
Cum evaluezi afirmațiile conform cărora există un număr mare de organisme care nu îmbătrânesc în natură, iar lista acestora crește de la an la an?
Din păcate, teoriile populare conform cărora există atât celule care nu sunt îmbătrânite, cât și organisme care nu sunt îmbătrânite, nu dispun de dovezi suficiente. Într-adevăr, în fiecare an, cercul animalelor „fără vârstă” se extinde inexorabil. La început acestea erau practic doar organisme unicelulare, apoi li s-au adăugat organisme multicelulare inferioare (hidra, moluște, arici de mare etc.). Și acum au apărut capete fierbinți care „descoperă” anumite specii fără vârstă chiar și printre pești, reptile și păsări. Așa va merge - în curând vor ajunge la mamifere și vor stabili, de exemplu, că nici elefanții nu îmbătrânesc, ci mor pur și simplu din cauza excesului de greutate corporală!
- Ești convins că nu există animale fără vârstă?
Nu sunt convins că nu există astfel de animale (deși sunt înclinat să o fac), dar că nu există nici o singură specie de animal pentru care absența îmbătrânirii să fi fost dovedită cu absolut încredere. În ceea ce privește celulele umane (precum și celulele și alți reprezentanți ai lumii animale), gradul de încredere este poate și mai mare: celulele stem, celulele germinale și chiar celulele tumorale, în principiu, vârsta. Celulele stem au fost considerate indiscutabil fără vârstă, dar acum apar lucrări experimentale care demonstrează contrariul.
- Pe ce se bazează această încredere? Ați efectuat singur experimentele relevante?
În general, cu foarte mult timp în urmă, în 1977-1980, am încercat să abordez problema îmbătrânirii în experimente pe șoareci. Dar rezultatele nu foarte fiabile (deși păreau să confirme ipoteza inițială) au convins că este mai bine să se facă analiză decât să experimenteze. Și iată unul dintre rezultatele acestei analize - conceptul de „Anerem” sau teoria ameiotică a îmbătrânirii. Include șase teze (postulate, dacă doriți), dintre care una (prima) este pur opera mea, iar restul sunt formulate pe baza unor idei deja existente în literatura de specialitate. Și, desigur, este important ca toate aceste teze să formeze o imagine destul de clară în ansamblu.
Deci, conceptul ameiotic, dacă este respectat, exclude posibilitatea existenței atât a celulelor neîmbătrânite în organismele multicelulare, cât și a organismelor neîmbătrânite (începând cu cele unicelulare). În același timp, desigur, sunt conștient că toate tezele conceptului sunt încă ipoteze. Dar ele par mult mai rezonabile decât alte opinii.
Deci, conceptul tău este ca un tester, cu ajutorul căruia poți evalua, relativ vorbind, adevărul anumitor presupuneri? În acest caz, spuneți-ne mai multe despre el.
Voi încerca să fac acest lucru cât mai accesibil posibil. Însuși numele conceptului („Anerem”) este o abreviere pentru cuvintele autocataliza, instabilitate, reparație, meioză. Teza unu. Îți amintești că definiția vieții lui Engels era anterior foarte bine cunoscută: „Viața este modul de existență al corpurilor proteice”? Am revizuit această definiție și am dat-o pe a mea, care a constituit prima teză: „Viața este o metodă de multiplicare autocatalitică a ADN-ului (mai puțin frecvent ARN) în natură”. Aceasta înseamnă că forța motrice din spatele atât apariției vieții, cât și a evoluției sale ulterioare este dorința de nesfârșit a acizilor nucleici de auto-reproducere fără sfârșit. În esență, orice organism este o biomașină îmbunătățită evolutiv, concepută pentru a păstra și multiplica eficient genomul pe care îl conține, urmată de distribuirea eficientă a copiilor sale în mediu.
- Este neobișnuit să te simți ca o biomașină...
Nimic, senzația va trece, dar funcția, scuzați-mă, va rămâne. Teza a doua: „Instabilitatea genomului este un element central al îmbătrânirii”. Exact așa înțeleg cei mai buni oameni de știință din Occident și aici, îmbătrânirea. Faptul este că, cu toate abilitățile lor remarcabile, acizii nucleici sunt susceptibili la efectele dăunătoare ale multor factori - radicali liberi, specii reactive de oxigen etc. Și deși în evoluție s-au creat multe sisteme de protecție (cum ar fi sistemul antioxidant), numeroase daune apar în mod constant în firele de ADN. Pentru a le detecta și corecta, există un alt sistem de protecție – repararea (restaurarea) ADN-ului. Următoarea teză, a treia, este un filtru care filtrează tot ce nu este „îmbătrânit”: „Repararea genomului în celulele mitotice și post-mitotice nu este completă”. Adică, orice sistem de reparare din aceste celule nu asigură corectarea de 100% a tuturor defectelor ADN care apar. Și asta înseamnă natura universală a îmbătrânirii.
- Dar dacă totul și toată lumea îmbătrânește, atunci cum se menține viața pe Pământ?
Ei bine, am devenit interesat de această problemă în 1977. Și am găsit, după cum mi s-a părut, răspunsul meu, deși zac la suprafață. Și 25 de ani mai târziu, în 2002, uitându-mă prin cărțile mele vechi, mi-am dat seama că această ipoteză nu era deloc a mea, dar citisem despre ea cu un an înainte în cartea lui M. M. Vilenchik, am uitat cu bucurie și apoi mi-am amintit, dar am perceput-o. ca a ta. Acestea sunt ciudateniile memoriei. Dar în cele din urmă, esența materiei este cea care contează, nu ambițiile descoperitorului.
Esența este formulată de a patra teză: „Repararea eficientă poate fi realizată numai în meioză (sau în versiunea sa simplificată - endomixis) - în timpul conjugării (fuziunii) cromozomilor.” Toată lumea pare să fi învățat ce este meioza la școală, dar, din păcate, uneori nici studenții noștri la medicină nu știu asta. Permiteți-mi să vă reamintesc: meioza este ultima diviziune dublă în formarea celulelor germinale - spermatozoizi și ovule. Apropo, vă spun un secret: femeile nu formează ouă. În ei, a doua diviziune meiotică (în stadiul de ovocit II - dezvoltarea celulei reproducătoare feminine) nu poate avea loc independent - fără ajutorul unui spermatozoid. Pentru că celula și-a „pierdut” centriolii (corpurile din celula implicate în diviziune) undeva: erau doar acolo (în timpul diviziunii anterioare), dar acum au plecat undeva. Iar fertilizarea ovocitului II este absolut necesară pentru ca spermatozoizii să-și aducă centriolii și să salveze situația. Văd asta ca „lucruri feminine” tipice. Deci a doua diviziune meiotică are loc în cele din urmă, dar celula rezultată nu mai este un ou, ci un zigot.
Ne-am lăsat duși de „lucruri feminine” și nu am clarificat cum se realizează repararea completă a ADN-ului în meioză.
Prima diviziune a meiozei este precedată de o profază foarte lungă: în gametogeneza masculină durează o lună întreagă, iar în gametogeneza feminină durează până la câteva decenii! În acest moment, cromozomii omologi se apropie unul de celălalt și rămân în această stare aproape tot timpul profazei.
În același timp, enzimele sunt puternic activate, tăind și cusând firele de ADN. Se credea că acest lucru era necesar doar pentru încrucișare - schimbul de cromozomi în secțiunile lor, ceea ce crește variabilitatea genetică a speciei. Într-adevăr, genele „tatălui” și „mamei”, care sunt încă distribuite în fiecare pereche de cromozomi omologi (identici din punct de vedere structural) pe diferiți cromozomi, se dovedesc a fi amestecate după trecere.
Dar M. M. Vilenchik, și după el eu, au atras atenția asupra faptului că enzimele de încrucișare sunt foarte asemănătoare cu enzimele de reparare a ADN-ului, în care, prin tăierea zonelor deteriorate, este, de asemenea, necesar să se rupă și să se coasă firele de ADN. Adică, super-repararea ADN-ului are loc probabil simultan cu trecerea. Ne putem imagina alte mecanisme de „reparare” majoră a genelor în timpul meiozei. Într-un fel sau altul, în acest caz, are loc o „întinerire” radicală (mai precis, completă) a celulelor, motiv pentru care celulele germinale mature încep să numere timpul ca de la zero. Dacă ceva nu funcționează, atunci senzorii de auto-monitorizare pentru starea propriului ADN sunt declanșați în celulă și începe procesul de apoptoză - auto-
uciderea celulei.
- Deci, în natură, întinerirea are loc doar în celulele germinale maturizate?
Absolut corect. Dar acest lucru este suficient pentru a asigura nemurirea speciei - pe fondul, vai, al mortalității inevitabile a tuturor indivizilor. La urma urmei, celulele sexuale sunt singurele! - singurul substrat material al organismelor părinte din care se naște o nouă viață - viața urmașilor.
Și faptul că acest mecanism se referă doar la celulele germinale este discutat în cele două teze rămase ale conceptului, care punctează toate i-urile. Teza cinci: „Meioza îmbunătățește starea genomului numai în generațiile ulterioare (mai multe generații deodată în organismele simple și doar una în toate celelalte).” Teza a șasea: „De aici urmează inevitabilitatea îmbătrânirii indivizilor (indivizilor) și nemurirea relativă a speciei în ansamblu.”
- Ce, meioza apare la toate speciile de animale?
Ar trebui să fie prezent la toate speciile de animale - conform conceptului Anerem, dacă se dovedește a fi corect. Într-adevăr, conceptul se bazează pe universalitatea nu numai a îmbătrânirii, ci și a meiozei. Am cercetat temeinic această problemă folosind datele din literatură. Desigur, la animalele suficient de dezvoltate - pești și cele „superioare” - există doar o metodă sexuală de reproducere, care implică și prezența meiozei. În plus, există sectoare uriașe atât ale florei, cât și ale faunei în care sunt comune tipuri mixte de reproducere. Aceasta înseamnă că alternează acte mai mult sau mai puțin prelungite de reproducere asexuată (de exemplu, diviziuni mitotice, sporulare, înmugurire, fragmentare etc.) și acte unice de reproducere sexuală sau cvasisexuală. O caracteristică esențială a procesului cvasisexual (așa-numitul endomix) este că și aici are loc o unire a cromozomilor identici din punct de vedere structural din seturile paternă și maternă (conjugarea cromozomilor omologi), deși nu se termină cu lor. divergenta in celule diferite.
Astfel, cu reproducerea mixtă, mai multe generații de organisme trăiesc, ca și cum ar îmbătrâni treptat (asemănător cu modul în care celulele care se divid mitotic îmbătrânesc la animalele mai complexe), apoi procesul sexual readuce organismele individuale la vârsta „zero” și oferă
oferă o viață confortabilă pentru mai multe generații. În cele din urmă, se crede că o serie de animale simple se reproduc doar asexuat. Dar în legătură cu ele, mai am ceva îndoieli: aceste organisme, într-o serie lungă de reproducere asexuată, nu au văzut ceva asemănător cu meioza sau endomixis (autofertilizarea)?
Se pare că conceptul pe care îl dezvoltați pune capăt tuturor viselor de extindere a vieții umane. La urma urmei, celulele obișnuite (nereproductive) sunt sortite să îmbătrânească și să îmbătrânească?
Nu, nu pun o cruce. În primul rând, pentru că ceea ce este mult mai important pentru noi nu este îmbătrânirea în sine, ci viteza acestui proces. Și puteți influența rata de îmbătrânire prin multe mijloace. Unele dintre ele sunt cunoscute, unii (cum ar fi ionii lui Skulachev) sunt în stadiul de cercetare, unii vor fi descoperiți mai târziu.
În al doilea rând, este posibil ca în timp să fie posibilă inițierea unor procese meiotice în celulele somatice - de exemplu, în celulele stem și nedivizoare. Mă refer la acele procese care restabilesc starea genomului: aceasta este aparent conjugarea cromozomilor omologi, crossing over, sau ceva mai subtil și încă necunoscut. Nu văd niciun motiv pentru care acest lucru ar fi imposibil în principiu. În liniile celulare germinale, meioza este pătrunsă de celule care, în general, au aceeași structură ca multe altele. Mai mult, chiar și după conjugarea cromozomilor, activitatea genelor corespunzătoare rămâne în acestea din urmă. Cu toate acestea, pentru a implementa acest proiect, este necesar să se identifice mai întâi genele responsabile pentru diferitele aspecte ale meiozei și să se stabilească modalități de a le viza. Acesta este, desigur, un proiect foarte fantastic. Cu toate acestea, mare parte din ceea ce avem astăzi nu părea fantastic ieri?!