Kod genetyczny zawiera. Biosynteza białek i kwasów nukleinowych
Wcześniej podkreślaliśmy, że nukleotydy mają cechę istotną dla tworzenia życia na Ziemi – jeśli w roztworze znajduje się jeden łańcuch polinukleotydowy, proces tworzenia drugiego (równoległego) łańcucha zachodzi samoistnie na zasadzie komplementarnego połączenia spokrewnionych nukleotydy. Warunkiem realizacji takich reakcji jest taka sama liczba nukleotydów w obu łańcuchach i ich związek chemiczny. Jednak podczas syntezy białek, gdy informacja z mRNA jest implementowana do struktury białka, nie może być mowy o przestrzeganiu zasady komplementarności. Wynika to z faktu, że nie tylko liczba monomerów jest różna w mRNA i w syntetyzowanym białku, ale także, co szczególnie ważne, nie ma między nimi podobieństwa strukturalnego (z jednej strony nukleotydy, z drugiej , aminokwasy). Jasne jest, że w tym przypadku konieczne staje się stworzenie nowej zasady dokładnej translacji informacji z polinukleotydu na strukturę polipeptydu. W ewolucji taka zasada została stworzona, a u jej podstaw ustanowiono kod genetyczny.
Kod genetyczny to system rejestrowania informacji dziedzicznych w cząsteczkach kwasu nukleinowego, oparty na pewnej przemianie sekwencji nukleotydów w DNA lub RNA, tworzących kodony odpowiadające aminokwasom w białku.
Kod genetyczny ma kilka właściwości.
Trójka.
Degeneracja lub nadmiarowość.
Jednoznaczność.
Biegunowość.
Bez nakładania się.
Ścisłość.
Wszechstronność.
Należy zauważyć, że niektórzy autorzy proponują także inne właściwości kodu związane z właściwościami chemicznymi nukleotydów zawartych w kodzie lub z częstością występowania poszczególnych aminokwasów w białkach organizmu itp. Jednak te właściwości wynikają z powyższego, więc rozważymy je tam.
a. Trójka. Kod genetyczny, podobnie jak wiele złożonych systemów, ma najmniejszą jednostkę strukturalną i funkcjonalną. Trójka to najmniejsza strukturalna jednostka kodu genetycznego. Składa się z trzech nukleotydów. Kodon jest najmniejszą funkcjonalną jednostką kodu genetycznego. Z reguły tryplety mRNA nazywane są kodonami. W kodzie genetycznym kodon pełni kilka funkcji. Po pierwsze, jego główną funkcją jest to, że koduje jeden aminokwas. Po drugie, kodon może nie kodować aminokwasu, ale w tym przypadku pełni inną funkcję (patrz niżej). Jak widać z definicji, tryplet to pojęcie charakteryzujące podstawowy jednostka strukturalna kod genetyczny (trzy nukleotydy). Kodon - charakteryzuje elementarna jednostka semantyczna genom – trzy nukleotydy warunkują przyłączenie jednego aminokwasu do łańcucha polipeptydowego.
Elementarną jednostkę strukturalną rozszyfrowano najpierw teoretycznie, a następnie potwierdzono eksperymentalnie jej istnienie. Rzeczywiście, 20 aminokwasów nie może być kodowanych przez jeden lub dwa nukleotydy. te ostatnie to tylko 4. Trzy z czterech nukleotydów dają 4 3 = 64 warianty, co więcej niż przekracza liczbę aminokwasów dostępnych w żywych organizmach (patrz Tabela 1).
Kombinacje nukleotydów przedstawione w Tabeli 64 mają dwie cechy. Po pierwsze, z 64 wariantów trojaczków tylko 61 to kodony i kodują dowolny aminokwas, nazywa się je kodony czuciowe... Trzy trojaczki nie kodują
Tabela 1.
Kodony Messenger RNA i odpowiadające im aminokwasy
|
B o n i ja c o d o n o v |
|||||
|
Nonsens |
Nonsens |
||||
|
Nonsens | |||||
|
Spotkał |
|||||
|
Wał |
|||||
aminokwasy a są sygnałami stop wskazującymi na koniec translacji. Są trzy takie trojaczki - UAA, UAG, UGA, są również nazywane „bezsensownymi” (kodonami nonsensownymi). W wyniku mutacji, która jest związana z wymianą jednego nukleotydu w trójce na inny, z kodonu sensownego może powstać kodon pozbawiony znaczenia. Ten rodzaj mutacji nazywa się nonsensowna mutacja... Jeżeli taki sygnał stop powstanie wewnątrz genu (w jego części informacyjnej), to podczas syntezy białka w tym miejscu proces będzie stale przerywany – syntetyzowana będzie tylko pierwsza (przed sygnałem stop) część białka. Osoba z tą patologią będzie miała brak białka i objawy związane z tym brakiem. Na przykład ten rodzaj mutacji został znaleziony w genie kodującym łańcuch beta hemoglobiny. Syntetyzowany jest skrócony nieaktywny łańcuch hemoglobiny, który szybko ulega zniszczeniu. W efekcie powstaje cząsteczka hemoglobiny pozbawiona łańcucha beta. Oczywiste jest, że taka cząsteczka prawdopodobnie nie spełni w pełni swoich obowiązków. Występuje poważna choroba, rozwijająca się w postaci niedokrwistości hemolitycznej (talasemia beta-zero, od greckiego słowa „Talas” – Morze Śródziemne, gdzie po raz pierwszy odkryto tę chorobę).
Mechanizm działania kodonów stop jest inny niż kodonów sensownych. Wynika to z faktu, że dla wszystkich kodonów kodujących aminokwasy znaleziono odpowiednie tRNA. Nie znaleziono tRNA dla nonsensownych kodonów. W konsekwencji tRNA nie bierze udziału w procesie zatrzymania syntezy białek.
KodonSIE (w bakteriach, czasem GUG) nie tylko kodują aminokwas metioninę i walinę, ale takżeinicjator transmisji .
b. Degeneracja lub nadmiarowość.
61 z 64 trojaczków koduje 20 aminokwasów. Takie trzykrotne przekroczenie liczby trypletów nad liczbą aminokwasów sugeruje, że w przekazie informacji można zastosować dwie opcje kodowania. Po pierwsze, nie wszystkie 64 kodony mogą być zaangażowane w kodowanie 20 aminokwasów, ale tylko 20, a po drugie, aminokwasy mogą być kodowane przez kilka kodonów. Badania wykazały, że natura skorzystała z tej drugiej opcji.
Jego preferencja jest oczywista. Gdyby tylko 20 z 64 wariantów trójek brało udział w kodowaniu aminokwasów, to 44 trójki (z 64) pozostałyby niekodujące, tj. bez znaczenia (bezsensowne kodony). Wcześniej zwracaliśmy uwagę na to, jak niebezpieczna dla życia komórki jest przekształcenie trójki kodującej w wyniku mutacji w kodon nonsensowny – to znacząco zaburza normalne działanie polimerazy RNA, prowadząc ostatecznie do rozwoju chorób. Obecnie w naszym genomie trzy kodony nie mają znaczenia, ale teraz wyobraź sobie, co by się stało, gdyby liczba nonsensownych kodonów wzrosła około 15 razy. Oczywiste jest, że w takiej sytuacji przejście od normalnych kodonów do nonsensownych kodonów będzie niezmiernie wyższe.
Kod, w którym jeden aminokwas jest kodowany przez kilka trypletów, nazywany jest zdegenerowanym lub nadmiarowym. Kilka kodonów odpowiada prawie każdemu aminokwasowi. Tak więc aminokwas leucyna może być kodowany przez sześć trypletów - UUA, UUG, CUU, CUTS, CUA, CUG. Walina jest kodowana przez cztery trojaczki, fenyloalanina przez dwie i tylko tryptofan i metionina są kodowane przez jeden kodon. Właściwość, która jest związana z zapisem tej samej informacji za pomocą różnych symboli, nazywa się degeneracja.
Liczba kodonów przypisanych do jednego aminokwasu dobrze koreluje z częstością występowania tego aminokwasu w białkach.
I to najprawdopodobniej nie jest przypadkowe. Im wyższa częstość występowania aminokwasu w białku, im częściej kodon tego aminokwasu jest prezentowany w genomie, tym większe prawdopodobieństwo jego uszkodzenia przez czynniki mutagenne. Dlatego jasne jest, że zmutowany kodon ma większe szanse na zakodowanie tego samego aminokwasu o wysokiej degeneracji. Z tych pozycji degeneracja kodu genetycznego jest mechanizmem chroniącym ludzki genom przed uszkodzeniem.
Należy zauważyć, że termin degeneracja jest używany w genetyce molekularnej iw innym sensie. Tak więc główna część informacji w kodonie przypada na pierwsze dwa nukleotydy, zasada w trzeciej pozycji kodonu okazuje się nieistotna. Zjawisko to nazywa się „degeneracją trzeciej bazy”. Ta ostatnia cecha minimalizuje efekt mutacji. Wiadomo na przykład, że główną funkcją czerwonych krwinek jest przenoszenie tlenu z płuc do tkanek i dwutlenku węgla z tkanek do płuc. Ta funkcja jest wykonywana przez pigment oddechowy - hemoglobinę, która wypełnia całą cytoplazmę erytrocytów. Składa się z części białkowej - globiny, która jest kodowana przez odpowiedni gen. Oprócz białka w cząsteczce hemoglobiny zawarty jest hem zawierający żelazo. Mutacje w genach globiny prowadzą do pojawienia się różnych wariantów hemoglobiny. Najczęściej mutacje są związane z zastąpienie jednego nukleotydu innym i pojawienie się nowego kodonu w genie, który może kodować nowy aminokwas w łańcuchu polipeptydowym hemoglobiny. W trójce w wyniku mutacji można zastąpić dowolny nukleotyd - pierwszy, drugi lub trzeci. Wiadomo, że kilkaset mutacji wpływa na integralność genów globiny. Blisko 400 z nich są związane z substytucją pojedynczych nukleotydów w genie i odpowiednią substytucją aminokwasową w polipeptydzie. Spośród nich tylko 100 substytucje prowadzą do niestabilności hemoglobiny i różnego rodzaju chorób od łagodnych do bardzo ciężkich. 300 (około 64%) mutacji substytucyjnych nie wpływa na czynność hemoglobiny i nie prowadzi do patologii. Jednym z powodów jest wspomniana wyżej „degeneracja trzeciej zasady”, kiedy zastąpienie trzeciego nukleotydu w triplecie kodującym serynę, leucynę, prolinę, argininę i niektóre inne aminokwasy prowadzi do pojawienia się kodonu synonimowego kodujący ten sam aminokwas. Fenotypowo ta mutacja się nie pojawi. Natomiast każda substytucja pierwszego lub drugiego nukleotydu w tryplecie w 100% przypadków prowadzi do pojawienia się nowego wariantu hemoglobiny. Ale nawet w tym przypadku może nie być poważnych zaburzeń fenotypowych. Powodem tego jest zastąpienie aminokwasu w hemoglobinie innym, podobnym do pierwszego pod względem właściwości fizykochemicznych. Na przykład, jeśli aminokwas o właściwościach hydrofilowych zostanie zastąpiony innym aminokwasem o tych samych właściwościach.
Hemoglobina składa się z grupy żelazowo-porfirynowej hemu (dołączają się do niej cząsteczki tlenu i dwutlenku węgla) oraz białka - globiny. Hemoglobina dorosłych (HbA) zawiera dwa identyczne -łańcuszki i dwa -więzy. Cząsteczka -łańcuch zawiera 141 reszt aminokwasowych, -łańcuch - 146, - oraz Łańcuchy β różnią się wieloma resztami aminokwasowymi. Sekwencja aminokwasowa każdego łańcucha globiny jest kodowana przez własny gen. Kodowanie genów - łańcuch znajduje się w krótkim ramieniu chromosomu 16, -gen - w krótkim ramieniu chromosomu 11. Substytucja w kodowaniu genu - łańcuch hemoglobiny pierwszego lub drugiego nukleotydu prawie zawsze prowadzi do pojawienia się nowych aminokwasów w białku, dysfunkcji hemoglobiny i poważnych konsekwencji dla pacjenta. Na przykład zastąpienie „C” w jednej z trójek CAU (histydyny) przez „Y” doprowadzi do pojawienia się nowej trójki CAU, która koduje inny aminokwas - tyrozynę. Łańcuch β polipeptydu histydyny do tyrozyny destabilizuje hemoglobinę. Choroba rozwija methemoglobinemię. Zastąpienie w wyniku mutacji kwasu glutaminowego waliną na 6 pozycji -łańcuchy są przyczyną najpoważniejszej choroby - anemii sierpowatej. Nie kontynuujmy smutnej listy. Zauważamy tylko, że po zastąpieniu dwóch pierwszych nukleotydów aminokwas może wydawać się podobny pod względem właściwości fizykochemicznych do poprzedniego. Tak więc zastąpienie drugiego nukleotydu w jednej z trójek kodujących kwas glutaminowy (GAA) w -łańcuch z „Y” prowadzi do pojawienia się nowego trypletu (GUA) kodującego walinę, a zastąpienie pierwszego nukleotydu przez „A” tworzy tryplet AAA kodujący aminokwas lizynę. Kwas glutaminowy i lizyna mają podobne właściwości fizykochemiczne - oba są hydrofilowe. Walina jest aminokwasem hydrofobowym. Dlatego zastąpienie hydrofilowego kwasu glutaminowego hydrofobową waliną znacząco zmienia właściwości hemoglobiny, co ostatecznie prowadzi do rozwoju anemii sierpowatokrwinkowej, natomiast zastąpienie hydrofilowego kwasu glutaminowego hydrofilową lizyną w mniejszym stopniu zmienia funkcję hemoglobiny – pacjenci mają postać łagodną anemii. W wyniku podstawienia trzeciej zasady nowa trójka może kodować te same aminokwasy, co poprzednia. Na przykład, jeśli uracyl został zastąpiony cytozyną w trójce CAC i pojawiła się trójka CAC, to praktycznie nie zostaną wykryte żadne zmiany fenotypowe u ludzi. Jest to zrozumiałe, ponieważ oba tryplety kodują ten sam aminokwas, histydynę.
Podsumowując, należy podkreślić, że degeneracja kodu genetycznego i degeneracja trzeciej podstawy z ogólnego biologicznego punktu widzenia są mechanizmami obronnymi osadzonymi w ewolucji w unikalnej strukturze DNA i RNA.
v. Jednoznaczność.
Każda trójka (oprócz pozbawionych znaczenia) koduje tylko jeden aminokwas. Zatem w kierunku kodon - aminokwas kod genetyczny jest jednoznaczny, w kierunku aminokwas - kodon jest niejednoznaczny (zdegenerowany).
Niedwuznaczny
Kodon aminokwasowy
Zdegenerowany
I w tym przypadku potrzeba jednoznaczności w kodzie genetycznym jest oczywista. W innym wariancie, podczas translacji tego samego kodonu, do łańcucha białkowego wprowadzane byłyby różne aminokwasy, w wyniku czego powstawałyby białka o różnych strukturach pierwszorzędowych i różnych funkcjach. Metabolizm komórkowy przestawiłby się na tryb działania „jeden gen – kilka poipeptydów”. Oczywiste jest, że w takiej sytuacji funkcja regulacyjna genów zostałaby całkowicie utracona.
Biegunowość
Odczytywanie informacji z DNA iz mRNA odbywa się tylko w jednym kierunku. Polaryzacja jest niezbędna do identyfikacji struktur wyższego rzędu (drugorzędne, trzeciorzędne itp.). Omówiliśmy wcześniej, że struktury niższego rzędu definiują struktury wyższego rzędu. Struktura trzeciorzędowa i struktury wyższego rzędu w białkach powstają natychmiast, gdy zsyntetyzowana nić RNA oddzieli się od cząsteczki DNA lub nić polipeptydowa odejdzie od rybosomu. Podczas gdy wolny koniec RNA lub polipeptydu uzyskuje strukturę trzeciorzędową, drugi koniec łańcucha jest nadal syntetyzowany na DNA (jeśli transkrybowany jest RNA) lub rybosomie (jeśli transkrybowany jest polipeptyd).
Dlatego jednokierunkowy proces odczytywania informacji (w syntezie RNA i białka) jest niezbędny nie tylko do określenia sekwencji nukleotydów lub aminokwasów w syntetyzowanej substancji, ale do sztywnego określenia drugorzędowych, trzeciorzędowych itp. Struktury.
e. Brak nakładania się.
Kod może się nakładać i nie nakładać. Większość organizmów nie ma nakładającego się kodu. W niektórych fagach znajduje się nakładający się kod.
Istotą kodu nienakładającego się jest to, że nukleotyd jednego kodonu nie może być jednocześnie nukleotydem innego kodonu. Gdyby kod się nakładał, to sekwencja siedmiu nukleotydów (GCCHCUG) mogłaby kodować nie dwa aminokwasy (alanina-alanina) (ryc. 33, A), jak w przypadku kodu nienakładającego się, ale trzy (jeśli jeden nukleotyd jest wspólny) (ryc. 33, B) lub pięć (jeśli dwa nukleotydy są wspólne) (zob. ryc. 33, C). W ostatnich dwóch przypadkach mutacja dowolnego nukleotydu doprowadziłaby do zakłócenia sekwencji dwóch, trzech itd. aminokwasy.
Stwierdzono jednak, że pojedyncza mutacja nukleotydowa zawsze zakłóca włączenie jednego aminokwasu do polipeptydu. Jest to istotny argument przemawiający za nienakładającym się kodem.
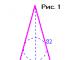
Wyjaśnijmy to na rysunku 34. Pogrubione linie pokazują tryplety kodujące aminokwasy w przypadku kodu nienakładającego się i nakładającego się. Eksperymenty jednoznacznie wykazały, że kod genetyczny się nie pokrywa. Nie wchodząc w szczegóły eksperymentu, zauważamy, że jeśli zastąpimy trzeci nukleotyd w sekwencji nukleotydowej (patrz ryc. 34)Posiadać (oznaczone gwiazdką) na coś innego:
1. W przypadku kodu nienakładającego się, białko kontrolowane przez tę sekwencję będzie miało podstawienie jednego (pierwszego) aminokwasu (oznaczonego gwiazdkami).
2. Przy nakładającym się kodzie w opcji A nastąpiłaby zmiana w dwóch (pierwszym i drugim) aminokwasach (oznaczonych gwiazdkami). W wariancie B zastąpienie wpłynęłoby na trzy aminokwasy (oznaczone gwiazdkami).
Jednak liczne eksperymenty wykazały, że gdy zaburzony jest jeden nukleotyd w DNA, zaburzenia w białku zawsze dotyczą tylko jednego aminokwasu, co jest charakterystyczne dla kodu nienakładającego się.
ГЦУГЦУГ ГЦУГЦУГ
ГЦУ ГЦУ ГЦУ УГЦ ЦУГ ГЦУ ЦУГ УГЦ ГЦУ ЦУГ
*** *** *** *** *** ***
Alanina - Alanin Ala - Cis - Lei Ala - Lei - Lei - Ala - Lei
A B C
Nienakładający się kod Nakładający się kod
Ryż. 34. Schemat wyjaśniający obecność nienakładającego się kodu w genomie (wyjaśnienie w tekście).
Brak nakładania się kodu genetycznego wiąże się z inną właściwością - odczyt informacji rozpoczyna się od pewnego punktu - sygnału inicjacji. Takim sygnałem inicjacji w mRNA jest kodon kodujący AUG metioniny.
Należy zauważyć, że ludzie nadal mają niewielką liczbę genów, które odbiegają od ogólnej zasady i nakładają się.
e. Zwartość.
Między kodonami nie ma znaków interpunkcyjnych. Innymi słowy, trojaczki nie są oddzielone od siebie, na przykład, jednym nic nie znaczącym nukleotydem. Brak „znaków interpunkcyjnych” w kodzie genetycznym udowodniono w eksperymentach.
F. Wszechstronność.
Kod jest taki sam dla wszystkich organizmów żyjących na Ziemi. Bezpośredni dowód uniwersalność kodu genetycznego uzyskano porównując sekwencje DNA z odpowiednimi sekwencjami białkowymi. Okazało się, że we wszystkich genomach bakteryjnych i eukariotycznych stosuje się te same zestawy wartości kodów. Są wyjątki, ale nie wiele.
Pierwsze wyjątki od uniwersalności kodu genetycznego znaleziono w mitochondriach niektórych gatunków zwierząt. Dotyczyło to kodonu terminatora UGA, który odczytywano w taki sam sposób, jak kodon UGG kodujący aminokwas tryptofan. Stwierdzono inne, rzadsze odstępstwa od uniwersalności.
MH. Kod genetyczny to system rejestrowania informacji dziedzicznych w cząsteczkach kwasu nukleinowego, oparty na pewnej przemianie sekwencji nukleotydów w DNA lub RNA, które tworzą kodony,
odpowiadające aminokwasom w białku.Kod genetyczny ma kilka właściwości.
W metabolizmie organizmu główna rola
należy do białek i kwasów nukleinowych.
Substancje białkowe stanowią podstawę wszystkich ważnych struktur komórkowych, mają niezwykle wysoką reaktywność i są wyposażone w funkcje katalityczne.
Kwasy nukleinowe wchodzą w skład najważniejszego organu komórki - jądra, jak również cytoplazmy, rybosomu, mitochondriów itp. Kwasy nukleinowe odgrywają ważną, nadrzędną rolę w dziedziczności, zmienności organizmu, w syntezie białek.
Plan synteza białko jest magazynowane w jądrze komórkowym, a synteza zachodzi bezpośrednio poza jądrem, dlatego jest konieczna Dostawa kodowane plan od jądra do miejsca syntezy. Ta usługa dostarczania jest realizowana przez cząsteczki RNA.
Proces rozpoczyna się o rdzeń komórki: część „drabiny” DNA rozwija się i otwiera. Dzięki temu litery RNA tworzą wiązania z otwarte listy DNA to jedna z nici DNA. Enzym przenosi litery RNA, aby połączyć je w nić. W ten sposób litery DNA są „przepisywane” na litery RNA. Nowo utworzona nić RNA zostaje odłączona, a „drabinka” DNA ponownie się zwija. Proces odczytywania informacji z DNA i syntezy ich z jego macierzy RNA nazywa się transkrypcja , a zsyntetyzowany RNA nazywa się informacyjnym lub i-RNA .
Po dalszych modyfikacjach taki kodowany i-RNA jest gotowy. i-RNA wychodzi z rdzenia i trafia do miejsca syntezy białek, gdzie dekodowane są litery i-RNA. Każdy zestaw trzech liter i-RNA tworzy „literę” reprezentującą jeden konkretny aminokwas.
Inny rodzaj RNA szuka tego aminokwasu, wychwytuje go za pomocą enzymu i dostarcza do miejsca syntezy białka. Ten RNA nazywa się transportowym RNA lub t-RNA. Gdy wiadomość i-RNA jest odczytywana i tłumaczona, łańcuch aminokwasów rośnie. Ten łańcuch skręca się i składa w unikalny kształt, tworząc jeden rodzaj białka. Na uwagę zasługuje nawet proces fałdowania białek: wszystko obliczyć za pomocą komputera opcje sfałdowanie średniej wielkości białka składającego się ze 100 aminokwasów zajęłoby 1027 (!) lat. A do utworzenia łańcucha 20 aminokwasów w organizmie nie trwa dłużej niż jedna sekunda, a proces ten zachodzi w sposób ciągły we wszystkich komórkach ciała.
Geny, kod genetyczny i jego właściwości.
Na Ziemi żyje około 7 miliardów ludzi. Z wyjątkiem 25-30 milionów par bliźniąt jednojajowych genetycznie wszyscy ludzie są różni : każdy jest wyjątkowy, posiada unikalne cechy dziedziczne, cechy charakteru, zdolności, temperament.
Takie różnice są wyjaśnione różnice w genotypach- zestawy genów organizmu; każdy jest wyjątkowy. Cechy genetyczne konkretnego organizmu są ucieleśnione w białkach - dlatego struktura białka jednej osoby różni się, choć dość nieznacznie, od białka innej osoby.
To nie znaczyże ludzie nie mają dokładnie tych samych białek. Białka pełniące te same funkcje mogą być takie same lub nieznacznie różnić się od siebie jednym lub dwoma aminokwasami. Ale nie istnieje na Ziemi ludzie (z wyjątkiem bliźniąt jednojajowych), którzy mieliby wszystkie białka są takie same .
Informacje o pierwotnej strukturze białka zakodowana jako sekwencja nukleotydów w regionie cząsteczki DNA, gen - jednostka informacji dziedzicznej organizmu. Każda cząsteczka DNA zawiera wiele genów. Całość wszystkich genów organizmu sprawia, że genotyp ... W ten sposób,
Gen to jednostka informacji dziedzicznej organizmu, która odpowiada oddzielnej sekcji DNA
Informacje dziedziczne są kodowane za pomocą kod genetyczny , który jest uniwersalny dla wszystkich organizmów i różni się jedynie naprzemianą nukleotydów tworzących geny i kodujących białka określonych organizmów.
Kod genetyczny składa się z trójek (trójek) nukleotydów DNA, połączonych w różnej sekwencji (AAT, HCA, ACG, THC itp.), z których każdy koduje określony aminokwas (który zostanie wstawiony do łańcucha polipeptydowego).
Tak właściwie kod
liczy się sekwencja nukleotydów w cząsteczce i-RNA
odkąd usuwa informacje z DNA (proces transkrypcje
) i przekłada ją na sekwencję aminokwasów w cząsteczkach syntetyzowanych białek (proces transmisje
).
Skład i-RNA obejmuje nukleotydy A-C-G-U, których trojaczki nazywane są kodony
: trójka DNA CGT na i-RNA stanie się trójką HCA, a trójka DNA AAG stanie się trójką UUC. Dokładnie tak kodony i-RNA
kod genetyczny jest odzwierciedlony w zapisie.
W ten sposób, kod genetyczny - ujednolicony system zapisu informacji dziedzicznej w cząsteczkach kwasu nukleinowego w postaci sekwencji nukleotydów ... Kod genetyczny opiera się na użyciu alfabetu składającego się tylko z czterech liter-nukleotydów, różniących się zasadami azotowymi: A, T, G, C.
Główne właściwości kodu genetycznego:

1. Kod genetyczny tryplet... Triplet (kodon) - sekwencja trzech nukleotydów, która koduje jeden aminokwas. Ponieważ białka zawierają 20 aminokwasów, oczywiste jest, że każdy z nich nie może być kodowany przez jeden nukleotyd ( ponieważ w DNA są tylko cztery rodzaje nukleotydów, to w tym przypadku 16 aminokwasów pozostaje niekodowanych). Brakuje również dwóch nukleotydów kodujących aminokwasy, ponieważ w tym przypadku można zakodować tylko 16 aminokwasów. Znaczy, najmniejsza liczba nukleotydy kodujące jeden aminokwas muszą mieć co najmniej trzy. W tym przypadku liczba możliwych trójek nukleotydów wynosi 43 = 64.
2. Redundancja (degeneracja) kod jest konsekwencją jego trypletowej natury i oznacza, że jeden aminokwas może być kodowany przez kilka trypletów (ponieważ jest 20 aminokwasów i 64 tryplety), z wyjątkiem metioniny i tryptofanu, które są kodowane tylko przez jeden tryplet. Ponadto niektóre tryplety pełnią określone funkcje: w cząsteczce i-RNA trójki UAA, UAG, UGA są kodonami terminacji, tj. zatrzymać-sygnały zatrzymujące syntezę łańcucha polipeptydowego. Trójka odpowiadająca metioninie (AUG), zlokalizowana na początku łańcucha DNA, nie koduje aminokwasu, ale pełni funkcję inicjacji (wzbudzenia) odczytu.
3. Jednoznaczność kod - wraz z redundancją kod ma właściwość jednoznaczność : każdy kodon pasuje tylko jeden specyficzny aminokwas.
4. Kolinearność kod, czyli sekwencja nukleotydów genu dokładnie odpowiada sekwencji aminokwasów w białku.
5. Kod genetyczny nie nakładające się i kompaktowe , czyli nie zawiera „znaków interpunkcyjnych”. Oznacza to, że proces odczytu nie dopuszcza możliwości nakładania się kolumn (trypletów), a począwszy od określonego kodonu, odczyt przebiega w sposób ciągły tryplet po tryplecie aż do zatrzymać-sygnały ( kodony terminacji).
6. Kod genetyczny wszechstronny to znaczy geny jądrowe wszystkich organizmów w ten sam sposób kodują informacje o białkach, niezależnie od poziomu organizacji i pozycji systemowej tych organizmów.
istnieje tabele kodów genetycznych do odszyfrowania kodony i-RNA i budujące łańcuchy cząsteczek białkowych.

Reakcje syntezy matrycy.
W żywych systemach zachodzą reakcje nieznane w przyrodzie nieożywionej - reakcje syntezy macierzy.
Termin „matryca” w technologii oznaczają formę używaną do odlewania monet, medali, typograficzny: utwardzony metal dokładnie odtwarza wszystkie szczegóły formy, z której wykonano odlew. Synteza macierzy przypomina odlewanie na matrycy: nowe molekuły są syntetyzowane ściśle według planu określonego w strukturze już istniejących molekuł.
Zasada macierzowa leży u źródła najważniejsze reakcje syntetyczne komórki, takie jak synteza kwasów nukleinowych i białek. Reakcje te zapewniają dokładną, ściśle określoną sekwencję jednostek monomerowych w zsyntetyzowanych polimerach.
W tym miejscu kierunkowy wciąganie monomerów w określone miejsce komórki - na cząsteczkach, które służą jako matryca, w której przebiega reakcja. Gdyby takie reakcje zachodziły w wyniku przypadkowego zderzenia cząsteczek, postępowałyby nieskończenie wolno. Synteza złożonych cząsteczek w oparciu o zasadę matrix jest szybka i dokładna. Rola matrycy makrocząsteczki kwasu nukleinowego biorą udział w reakcjach macierzowych DNA lub RNA .

Cząsteczki monomeryczne z którego syntetyzowany jest polimer – nukleotydy lub aminokwasy – zgodnie z zasadą komplementarności są umiejscowione i utrwalone na matrycy w ściśle określonej, określonej kolejności.
Potem się dzieje „sieciowanie” jednostek monomeru w łańcuch polimerowy a gotowy polimer usuwa się z matrycy.
Odtąd matryca jest gotowa do montażu nowej cząsteczki polimeru. Oczywiste jest, że tak jak na danej formie można rzucić tylko jedną monetę, jedną literę, tak na danej cząsteczce matrycy można „zmontować” tylko jeden polimer.
Macierzowy typ reakcji- specyficzna cecha chemii systemów żywych. Są podstawą podstawowej właściwości wszystkich żywych istot - ich zdolności do reprodukcji własnego gatunku.
Reakcje syntezy matrycy
1. replikacja DNA - replikacja (z łac. repliktio - odnowienie) - proces syntezy cząsteczki potomnej kwasu dezoksyrybonukleinowego na macierzy macierzystej cząsteczki DNA. Podczas późniejszego podziału komórki macierzystej każda komórka potomna otrzymuje jedną kopię cząsteczki DNA, która jest identyczna z DNA pierwotnej komórki macierzystej. Proces ten zapewnia dokładną transmisję informacji genetycznej z pokolenia na pokolenie. Replikacja DNA jest przeprowadzana przez złożony kompleks enzymatyczny składający się z 15-20 różnych białek, zwanych replikasoma ... Materiałem do syntezy są wolne nukleotydy obecne w cytoplazmie komórek. Biologiczne znaczenie replikacji polega na dokładnym przekazywaniu informacji dziedzicznych z cząsteczki rodzicielskiej na cząsteczkę potomną, co zwykle ma miejsce podczas podziału komórki somatyczne.
Cząsteczka DNA składa się z dwóch komplementarnych nici. Te łańcuchy są słabe wiązania wodorowe które mogą zostać złamane przez enzymy. Cząsteczka DNA jest zdolna do samopodwojenia (replikacji), a nowa połowa cząsteczki jest syntetyzowana na każdej starej połowie cząsteczki.
Ponadto na cząsteczce DNA można zsyntetyzować cząsteczkę i-RNA, która następnie przenosi informacje otrzymane z DNA do miejsca syntezy białek.
Transfer informacji i synteza białek opierają się na zasadzie matrycy, porównywalnej z pracą prasy drukarskiej w drukarni. Informacje z DNA są kopiowane wielokrotnie. Jeśli błędy wystąpią podczas kopiowania, będą się powtarzać we wszystkich kolejnych kopiach.

To prawda, że niektóre błędy podczas kopiowania informacji przez cząsteczkę DNA można poprawić - nazywa się to procesem eliminowania błędów remont... Pierwszą z reakcji w procesie przekazywania informacji jest replikacja cząsteczki DNA i synteza nowych nici DNA.
2. Transkrypcja (z łac. transkrypcja – przepisywanie) – proces syntezy RNA z wykorzystaniem DNA jako matrycy, który zachodzi we wszystkich żywych komórkach. Innymi słowy, jest to transfer informacji genetycznej z DNA do RNA.
Transkrypcja jest katalizowana przez enzym polimerazę RNA zależną od DNA. Polimeraza RNA porusza się wzdłuż cząsteczki DNA w kierunku 3 „→ 5”. Transkrypcja składa się z etapów inicjacja, wydłużenie i zakończenie ... Jednostką transkrypcji jest operon, fragment cząsteczki DNA składający się z promotor, część transkrybowana i terminator ... i-RNA składa się z jednej nici i jest syntetyzowany na DNA zgodnie z zasadą komplementarności z udziałem enzymu, który aktywuje początek i koniec syntezy cząsteczki i-RNA.
Gotowa cząsteczka i-RNA wchodzi do cytoplazmy na rybosomy, gdzie zachodzi synteza łańcuchów polipeptydowych.
3. Audycja (od łac. tłumaczenie- transfer, ruch) – proces syntezy białek z aminokwasów na matrycy informacyjnego (macierzy) RNA (mRNA, mRNA), realizowany przez rybosom. Innymi słowy, jest to proces translacji informacji zawartej w sekwencji nukleotydowej m-RNA na sekwencję aminokwasów w polipeptydzie.

4. Transkrypcja odwrotna to proces tworzenia dwuniciowego DNA na podstawie informacji z jednoniciowego RNA. Proces ten nazywa się odwrotną transkrypcją, ponieważ w tym przypadku przekazywanie informacji genetycznej odbywa się w kierunku „odwrotnym”, w stosunku do transkrypcji. Idea odwrotnej transkrypcji była początkowo bardzo niepopularna, ponieważ była sprzeczna z centralnym dogmatem. Biologia molekularna, co sugerowało, że DNA ulega transkrypcji do RNA, a następnie translacji do białek.
 Jednak w 1970 roku Temin i Baltimore niezależnie odkryli enzym zwany odwrotna transkryptaza (odwrotna transkryptaza)
i ostatecznie potwierdzono możliwość odwrotnej transkrypcji. W 1975 roku Temin i Baltimore zostali nagrodzeni nagroda Nobla z zakresu fizjologii i medycyny. Niektóre wirusy (takie jak ludzki wirus niedoboru odporności, który powoduje zakażenie HIV) mają zdolność transkrypcji RNA do DNA. HIV ma genom RNA osadzony w DNA. W rezultacie DNA wirusa można połączyć z genomem komórki gospodarza. Główny enzym odpowiedzialny za syntezę DNA z RNA nazywa się odwróć... Jedną z funkcji rewersji jest tworzenie komplementarne DNA
(cDNA) z genomu wirusa. Powiązany enzym rybonukleaza tnie RNA, podczas gdy odwrotna transkryptaza syntetyzuje cDNA z podwójnej helisy DNA. cDNA jest integrowany z genomem komórki gospodarza za pomocą integrazy. Wynik to synteza białek wirusowych przez komórkę gospodarza które tworzą nowe wirusy. W przypadku HIV programowana jest również apoptoza (śmierć komórki) limfocytów T. W innych przypadkach komórka może pozostać dystrybutorem wirusów.
Jednak w 1970 roku Temin i Baltimore niezależnie odkryli enzym zwany odwrotna transkryptaza (odwrotna transkryptaza)
i ostatecznie potwierdzono możliwość odwrotnej transkrypcji. W 1975 roku Temin i Baltimore zostali nagrodzeni nagroda Nobla z zakresu fizjologii i medycyny. Niektóre wirusy (takie jak ludzki wirus niedoboru odporności, który powoduje zakażenie HIV) mają zdolność transkrypcji RNA do DNA. HIV ma genom RNA osadzony w DNA. W rezultacie DNA wirusa można połączyć z genomem komórki gospodarza. Główny enzym odpowiedzialny za syntezę DNA z RNA nazywa się odwróć... Jedną z funkcji rewersji jest tworzenie komplementarne DNA
(cDNA) z genomu wirusa. Powiązany enzym rybonukleaza tnie RNA, podczas gdy odwrotna transkryptaza syntetyzuje cDNA z podwójnej helisy DNA. cDNA jest integrowany z genomem komórki gospodarza za pomocą integrazy. Wynik to synteza białek wirusowych przez komórkę gospodarza które tworzą nowe wirusy. W przypadku HIV programowana jest również apoptoza (śmierć komórki) limfocytów T. W innych przypadkach komórka może pozostać dystrybutorem wirusów.
Sekwencję reakcji matrycowych w biosyntezie białek można przedstawić na diagramie.

W ten sposób, biosynteza białek- To jeden z rodzajów metabolizmu tworzyw sztucznych, podczas którego dziedziczna informacja zakodowana w genach DNA jest realizowana w określonej sekwencji aminokwasów w cząsteczkach białek.
Cząsteczki białka są zasadniczo łańcuchy polipeptydowe składa się z poszczególnych aminokwasów. Ale aminokwasy nie są wystarczająco aktywne, aby wiązać się same. Dlatego przed połączeniem się ze sobą i utworzeniem cząsteczki białka aminokwasy muszą: Aktywuj ... Ta aktywacja następuje pod działaniem specjalnych enzymów.
W wyniku aktywacji aminokwas staje się bardziej labilny i pod wpływem tego samego enzymu wiąże się z t- RNA... Każdy aminokwas odpowiada ściśle określonemu t- RNA, który znajduje „swój” aminokwas i przenosi się ją do rybosomu.
Dlatego różne aktywowane aminokwasy w połączeniu z ich T- RNA... Rybosom jest niejako przenośnik złożyć łańcuch białkowy z różnych wchodzących do niego aminokwasów.
Równolegle z t-RNA, na którym „siedzi” jego własny aminokwas, rybosom otrzymuje „ sygnał„Z DNA zawartego w jądrze. Zgodnie z tym sygnałem w rybosomie syntetyzowane jest określone białko.
Kierujący wpływ DNA na syntezę białek nie odbywa się bezpośrednio, ale za pomocą specjalnego mediatora - matryca lub informacyjne RNA (m-RNA lub i-RNA), który zsyntetyzowany w jądrze e pod wpływem DNA, dlatego jego skład odzwierciedla skład DNA. Cząsteczka RNA jest jak forma formy DNA. Zsyntetyzowany i-RNA wchodzi do rybosomu i niejako przenosi się do tej struktury plan- w jakiej kolejności należy łączyć ze sobą aktywowane aminokwasy wchodzące do rybosomu, aby zsyntetyzować określone białko. W przeciwnym razie, Informacja genetyczna zakodowany w DNA jest przenoszony na m-RNA, a następnie na białko.
Cząsteczka i-RNA wchodzi do rybosomu i szwy jej. Segment, który jest w ten moment w rybosomie, zdefiniowany kodon (tryplet), współdziała dość specyficznie z odpowiednią strukturą tryplet (antykodon) w transportowym RNA, który wprowadził aminokwas do rybosomu.
Transportowy RNA wraz z aminokwasem jest dopasowany do określonego kodonu mRNA i łączy z nim; do następnej, sąsiedniej strony i-RNA łączy inny t-RNA z innym aminokwasem i tak dalej, aż zostanie odczytany cały łańcuch i-RNA, aż wszystkie aminokwasy zostaną nawleczone w odpowiedniej kolejności, tworząc cząsteczkę białka. I t-RNA, który dostarcza aminokwas w określone miejsce łańcucha polipeptydowego, uwolniony od aminokwasu i pozostawia rybosom.
Następnie ponownie w cytoplazmie można do niej dołączyć wymagany aminokwas, który ponownie przeniesie go do rybosomu. W procesie syntezy białek zaangażowanych jest jednocześnie nie jeden, ale kilka rybosomów - polirybosomów.
Główne etapy transferu informacji genetycznej:
1. Synteza na DNA jak na matrycy i-RNA (transkrypcja)
2. Synteza w rybosomach łańcucha polipeptydowego zgodnie z programem zawartym w m-RNA (tłumaczenie)
.
Etapy są uniwersalne dla wszystkich żywych istot, ale czasowe i przestrzenne relacje tych procesów różnią się u pro- i eukariontów.
Posiadać prokariota transkrypcję i translację można przeprowadzić jednocześnie, ponieważ DNA znajduje się w cytoplazmie. Posiadać eukarionty transkrypcja i translacja są ściśle rozdzielone w czasie i przestrzeni: synteza różnych RNA zachodzi w jądrze, po czym cząsteczki RNA muszą opuścić jądro, przechodząc przez błonę jądrową. Następnie w cytoplazmie RNA są transportowane do miejsca syntezy białek.
Wykład 5. Kod genetyczny
Definicja pojęcia
Kod genetyczny to system zapisu informacji o sekwencji aminokwasów w białkach za pomocą sekwencji ułożenia nukleotydów w DNA.
Ponieważ DNA nie jest bezpośrednio zaangażowany w syntezę białek, kod jest napisany w języku RNA. RNA zawiera uracyl zamiast tyminy.
Właściwości kodu genetycznego
1. Trójka
Każdy aminokwas jest kodowany jako sekwencja 3 nukleotydów.
Definicja: tryplet lub kodon - sekwencja trzech nukleotydów, która koduje jeden aminokwas.
Kod nie może być singletowy, ponieważ 4 (liczba różnych nukleotydów w DNA) jest mniejsza niż 20. Kod nie może być dublet, ponieważ 16 (liczba kombinacji i permutacji 4 nukleotydów przez 2) jest mniejsza niż 20. Kod może być tryplet, ponieważ 64 (liczba kombinacji i permutacji od 4 do 3) to więcej niż 20.
2. Degeneracja.
Wszystkie aminokwasy, z wyjątkiem metioniny i tryptofanu, są kodowane przez więcej niż jeden tryplet:
2 AK 1 tryplet = 2.
9 AK 2 trojaczki = 18.
1 AK 3 trojaczki = 3.
5 AK 4 trojaczki = 20.
3 AK 6 trojaczków = 18.
Łącznie 61 trojaczków koduje 20 aminokwasów.
3. Obecność międzygenowych znaków interpunkcyjnych.
Definicja:
Gen to fragment DNA, który koduje jeden łańcuch polipeptydowy lub jedną cząsteczkę tPHK, rRNA lubsPHK.
GenytPHK, rPHK, sPHKbiałka nie kodują.
Na końcu każdego genu kodującego polipeptyd znajduje się co najmniej jeden z 3 trypletów kodujących kodony stop lub sygnały stop RNA. W mRNA wyglądają tak: UAA, UAG, UGA ... Kończą (kończą) transmisję.
Konwencjonalnie kodon odnosi się również do znaków interpunkcyjnych SIE - pierwszy po sekwencji liderowej. (Patrz Wykład 8) Funkcjonuje jako wielka litera. W tej pozycji koduje formylometioninę (w prokariotach).
4. Jednoznaczność.
Każdy tryplet koduje tylko jeden aminokwas lub jest terminatorem translacji.
Wyjątkiem jest kodon SIE ... U prokariontów w pierwszej pozycji (duża litera) koduje formylometioninę, aw każdej innej metioninę.
5. Zwartość lub brak wewnątrzgenowych znaków interpunkcyjnych.
W genie każdy nukleotyd jest częścią kodonu znaczenia.
W 1961 Seymour Benzer i Francis Crick udowodnili eksperymentalnie, że kod jest trójkowy i zwarty.
Istota eksperymentu: mutacja „+” – wstawienie jednego nukleotydu. Mutacja „-” to utrata jednego nukleotydu. Pojedyncza mutacja „+” lub „-” na początku genu psuje cały gen. Podwójna mutacja „+” lub „-” również psuje cały gen.
Potrójna mutacja „+” lub „-” na początku genu psuje tylko jego część. Poczwórna mutacja + lub - ponownie psuje cały gen.
Eksperyment udowadnia, że kod jest skomplikowany i wewnątrz genu nie ma znaków interpunkcyjnych. Eksperyment przeprowadzono na dwóch sąsiednich genach fagowych i wykazał dodatkowo obecność znaków interpunkcyjnych między genami.
6. Wszechstronność.
Kod genetyczny jest taki sam dla wszystkich stworzeń żyjących na Ziemi.
W 1979 roku otwarto Burrell ideał ludzki kod mitochondrialny.
Definicja:
„Idealny” to kod genetyczny, w którym spełniona jest zasada degeneracji kodu quasi-dubletu: jeśli pierwsze dwa nukleotydy pokrywają się w dwóch trypletach, a trzeci nukleotydy należą do tej samej klasy (obie są purynami lub oba są pirymidynami) , to te trojaczki kodują ten sam aminokwas...
W kodzie ogólnym istnieją dwa wyjątki od tej reguły. Oba odstępstwa od idealnego kodu w uniwersalności dotyczą fundamentalnych punktów: początku i końca syntezy białek:
Kodon | uniwersalny kod | Kody mitochondrialne |
|||
Kręgowce | Bezkręgowce | Drożdże | Rośliny |
||
ZATRZYMAĆ | ZATRZYMAĆ |
||||
Z UA | |||||
A G A | ZATRZYMAĆ | ||||
ZATRZYMAĆ | 230 podstawień nie zmienia klasy kodowanego aminokwasu. do zdolności rozdzierania. W 1956 Georgy Gamow zaproponował wariant nakładającego się kodu. Zgodnie z kodem Gamowa, każdy nukleotyd, począwszy od trzeciego w genie, zawiera się w 3 kodonach. Po rozszyfrowaniu kodu genetycznego okazało się, że nie nakładał się, tj. każdy nukleotyd jest zawarty tylko w jednym kodonie. Zalety nakładającego się kodu genetycznego: zwartość, mniejsza zależność struktury białka od insercji lub delecji nukleotydów. Wada: duża zależność struktury białka od substytucji nukleotydów i restrykcji na sąsiadach. W 1976 zsekwencjonowano DNA faga φX174. Ma jednoniciowy kolisty DNA złożony z 5375 nukleotydów. Wiadomo było, że fag koduje 9 białek. W przypadku 6 z nich zidentyfikowano geny zlokalizowane jeden po drugim. Okazało się, że się nakładają. Gen E jest całkowicie w obrębie genu D ... Jego kodon inicjacyjny pojawia się w wyniku przesunięcia odczytu o jeden nukleotyd. Gen J zaczyna się tam, gdzie kończy się gen D ... Kodon inicjacji genu J pokrywa się z kodonem terminacji genu D w wyniku przesunięcia dwóch nukleotydów. Konstrukt jest nazywany „przesunięciem ramki odczytu” przez liczbę nukleotydów, która nie jest wielokrotnością trzech. Do tej pory nakładanie się zostało wykazane tylko dla kilku fagów. Pojemność informacji DNA Na Ziemi żyje 6 miliardów ludzi. Informacje dziedziczne na ich temat 4x10 13 stron książki. Strony te zajmowałyby objętość 6 budynków NSU. 6x10 9 plemników zajmuje połowę naparstka. Ich DNA zajmuje mniej niż jedną czwartą naparstka. | ||||
- ujednolicony system rejestrowania informacji dziedzicznych w cząsteczkach kwasu nukleinowego w postaci sekwencji nukleotydów. Kod genetyczny opiera się na użyciu alfabetu składającego się tylko z czterech liter-nukleotydów, różniących się zasadami azotowymi: A, T, G, C.
Główne właściwości kodu genetycznego są następujące:
1. Kod genetyczny jest trójką. Triplet (kodon) - sekwencja trzech nukleotydów, która koduje jeden aminokwas. Ponieważ białka zawierają 20 aminokwasów, oczywiste jest, że każdy z nich nie może być kodowany przez jeden nukleotyd (ponieważ w DNA są tylko cztery rodzaje nukleotydów, w tym przypadku 16 aminokwasów pozostaje niezakodowanych). Brakuje również dwóch nukleotydów kodujących aminokwasy, ponieważ w tym przypadku można zakodować tylko 16 aminokwasów. Oznacza to, że najmniejsza liczba nukleotydów kodujących jeden aminokwas okazuje się być trzema. (W tym przypadku liczba możliwych trypletów nukleotydów wynosi 4 3 = 64).
2. Nadmiarowość (degeneracja) kodu jest konsekwencją jego trypletu i oznacza, że jeden aminokwas może być kodowany przez kilka trypletów (ponieważ jest 20 aminokwasów i 64 tryplety). Wyjątkiem są metionina i tryptofan, które są kodowane tylko przez jeden tryplet. Ponadto niektóre trojaczki mają określone funkcje. Tak więc w cząsteczce mRNA trzy z nich, UAA, UAH i UGA, są kodonami terminacji, to znaczy sygnałami zatrzymującymi, które zatrzymują syntezę łańcucha polipeptydowego. Trójka odpowiadająca metioninie (AUG), zlokalizowana na początku łańcucha DNA, nie koduje aminokwasu, ale pełni funkcję inicjacji (wzbudzenia) odczytu.
3. Jednocześnie z redundancją kod ma właściwość jednoznaczności, co oznacza, że każdy kodon odpowiada tylko jednemu określonemu aminokwasowi.
4. Kod jest współliniowy; sekwencja nukleotydów w genie dokładnie odpowiada sekwencji aminokwasów w białku.
5. Kod genetyczny jest niezachodzący na siebie i zwarty, to znaczy nie zawiera „znaków interpunkcyjnych”. Oznacza to, że proces odczytu nie dopuszcza możliwości nakładania się kolumn (trypletów) i rozpoczynając od określonego kodonu, odczyt przebiega w sposób ciągły, tryplet po tryplecie, aż do sygnałów zatrzymania (kodonów terminacji). Np. w mRNA następująca sekwencja zasad azotowych AUGGUGTSUUAAUGUG będzie odczytywana tylko przez takie trojaczki: AUG, GUG, CUU, AAU, GUG, a nie AUG, UGG, GGU, GUG itd. lub AUG, GGU, UGC, CUU itp. lub w inny sposób (na przykład kodon AUG, znak interpunkcyjny G, kodon UGC, znak interpunkcyjny U itp.).
6. Kod genetyczny jest uniwersalny, to znaczy geny jądrowe wszystkich organizmów kodują informacje o białkach w ten sam sposób, niezależnie od poziomu organizacji i pozycji systemowej tych organizmów.
Kod genetyczny- ujednolicony system rejestrowania informacji dziedzicznych w cząsteczkach kwasu nukleinowego w postaci sekwencji nukleotydów. Kod genetyczny opiera się na użyciu alfabetu składającego się tylko z czterech liter A, T, C, G, odpowiadających nukleotydom DNA. W sumie istnieje 20 rodzajów aminokwasów. Spośród 64 kodonów trzy - UAA, UAG, UGA - nie kodują aminokwasów, nazwano je kodonami nonsensownymi i pełnią funkcję znaków interpunkcyjnych. Kodon (kodujący trinukleotyd) to jednostka kodu genetycznego, trójka reszt nukleotydowych (tryplet) w DNA lub RNA, kodująca włączenie jednego aminokwasu. Same geny nie biorą udziału w syntezie białek. Mediatorem między genem a białkiem jest mRNA. Struktura kodu genetycznego charakteryzuje się tym, że jest tryplet, czyli składa się z trypletów (tryplet) zasad azotowych DNA, zwanych kodonami. 64
Właściwości genów. kod
1) Triplet: jeden aminokwas jest kodowany przez trzy nukleotydy. Te 3 nukleotydy w DNA
zwany triplet, w mRNA - kodon, w tRNA - antykodon.
2) Redundancja (degeneracja): jest tylko 20 aminokwasów, a trójki kodujące aminokwasy 61, dlatego każdy aminokwas jest kodowany przez kilka trójek.
3) Jednoznaczność: każdy tryplet (kodon) koduje tylko jeden aminokwas.
4) Wszechstronność: kod genetyczny jest taki sam dla wszystkich żywych organizmów na Ziemi.
5.) ciągłość i spójność kodonów podczas czytania. Oznacza to, że sekwencja nukleotydów jest odczytywana trójka po trójce bez przerw, podczas gdy sąsiednie trójki nie nakładają się.
88. Dziedziczność i zmienność to podstawowe właściwości istot żywych. Rozumienie Darwina zjawisk dziedziczności i zmienności.
Dziedziczność nazywają wspólną właściwością wszystkich organizmów zachowanie i przekazywanie cech od rodzica potomstwu. Dziedziczność- jest to właściwość organizmów do reprodukcji z pokolenia na pokolenie podobnego typu metabolizmu, który rozwinął się w tym procesie rozwój historyczny gatunek i przejawia się w określonych warunkach otoczenie zewnętrzne.
Zmienność zachodzi proces powstawania różnic jakościowych między osobnikami tego samego gatunku, który wyraża się albo zmianą pod wpływem środowiska zewnętrznego tylko jednego fenotypu, albo genetycznie uwarunkowanymi zmiennościami dziedzicznymi wynikającymi z kombinacji, rekombinacji i mutacji, które występują w wielu kolejnych pokoleniach i populacjach.
Rozumienie Darwina dziedziczności i zmienności.
Pod dziedziczeniem Darwin rozumiał zdolność organizmów do zachowania swojego gatunku, cech odmianowych i indywidualnych u potomstwa. Ta cecha była dobrze znana i stanowiła odmianę dziedziczną. Darwin szczegółowo przeanalizował znaczenie dziedziczności w procesie ewolucyjnym. Zwrócił uwagę na przypadki jednorodności mieszańców pierwszego pokolenia i rozszczepienia cech w drugim pokoleniu, był świadomy dziedziczności związanej z płcią, atawizmów hybrydowych i szeregu innych zjawisk dziedziczności.
Zmienność. Porównując wiele ras zwierząt i odmian roślin, Darwin zauważył, że w każdym gatunku zwierząt i roślin oraz w kulturze w obrębie jakiejkolwiek odmiany i rasy nie ma identycznych osobników. Darwin doszedł do wniosku, że zmienność jest nieodłączną cechą wszystkich zwierząt i roślin.
Analizując materiał dotyczący zmienności zwierząt, naukowiec zauważył, że każda zmiana warunków przetrzymywania wystarczy, aby spowodować zmienność. Tak więc Darwin rozumiał zmienność jako zdolność organizmów do nabywania nowych cech pod wpływem warunków środowiskowych. Wyróżnił następujące formy zmienności:
Specyficzna (grupowa) zmienność(teraz nazywany modyfikacja) - podobna zmiana we wszystkich osobnikach potomstwa w jednym kierunku ze względu na wpływ pewnych warunków. Niektóre zmiany są zwykle niedziedziczne.
Niepewna zmienność indywidualna(teraz nazywany genotypowy) - pojawienie się różnych nieznacznych różnic u osobników tego samego gatunku, odmiany, rasy, dzięki którym, istniejąc w podobnych warunkach, jedna osoba różni się od innych. Taka wielokierunkowa zmienność jest konsekwencją niepewnego wpływu warunków egzystencji na każdą jednostkę.
Współzależny(lub względna) zmienność. Darwin rozumiał organizm jako integralny system, którego poszczególne części są ze sobą ściśle powiązane. Dlatego zmiana struktury lub funkcji jednej części często powoduje zmianę w innej lub innych. Przykładem takiej zmienności jest związek między rozwojem funkcjonującego mięśnia a powstaniem grzebienia na kości, do której się przyczepia. U wielu ptaków brodzących istnieje korelacja między długością szyi a długością kończyn: ptaki z długą szyją mają również długie kończyny.
Zmienność kompensacyjna polega na tym, że rozwój jednych narządów lub funkcji jest często przyczyną ucisku innych, czyli istnieje odwrotna korelacja np. między wydajnością mleczną a mięsnością bydła.
89. Zmienność modyfikacji. Szybkość reakcji cech genetycznie uwarunkowanych. Fenokopie.
Fenotypowy zmienność obejmuje zmiany stanu bezpośrednich znaków, które zachodzą pod wpływem warunków rozwojowych lub czynników środowiskowych. Zakres zmienności modyfikacji jest ograniczony odpowiedzią normalną. Wynikająca z tego specyficzna zmiana modyfikacji cechy nie jest dziedziczona, ale zakres zmienności modyfikacji jest określany przez dziedziczność.W tym przypadku materiał dziedziczny nie jest zaangażowany w zmianę.
Szybkość reakcji- jest to granica zmienności modyfikacji cechy. Szybkość reakcji jest dziedziczona, ale nie same modyfikacje, tj. zdolność do rozwijania cechy, a forma jej manifestacji zależy od warunków środowiskowych. Szybkość reakcji jest specyficzną cechą ilościową i jakościową genotypu. Są znaki o dużej szybkości reakcji, wąskiej () i jednoznacznej szybkości. Szybkość reakcji ma limity lub granice dla każdego gatunku (dolne i górne) - na przykład zwiększone karmienie doprowadzi do wzrostu masy zwierzęcia, ale będzie mieściło się w szybkości reakcji charakterystycznej dla tego gatunku lub rasy. Szybkość reakcji jest uwarunkowana genetycznie i dziedziczona. Dla różnych znaków granice normy reakcji są bardzo różne. Na przykład wydajność mleczna, produktywność zbóż i wiele innych cech ilościowych ma szerokie granice szybkości reakcji, wąskie to intensywność barwy większości zwierząt i wiele innych cech jakościowych. Pod wpływem niektórych szkodliwe czynniki, z którym człowiek nie ma do czynienia w procesie ewolucji, wyklucza się możliwość zmienności modyfikacji, która determinuje normy reakcji.
Fenokopie- zmiany w fenotypie pod wpływem niekorzystnych czynników środowiskowych, objawiające się podobnie do mutacji. Powstałe modyfikacje fenotypowe nie są dziedziczone. Ustalono, że występowanie fenokopii jest związane z wpływem warunków zewnętrznych na pewien ograniczony etap rozwoju. Co więcej, ten sam czynnik, w zależności od fazy, na którą działa, może kopiować różne mutacje lub jeden etap reaguje na jeden czynnik, a drugi na inny. Do wywołania tej samej fenokopii można użyć różnych środków, co wskazuje, że nie ma związku między wynikiem zmiany a czynnikiem wpływającym. Najbardziej złożone genetyczne zaburzenia rozwojowe są stosunkowo łatwe do odtworzenia, podczas gdy cechy są znacznie trudniejsze do skopiowania.
90. Adaptacyjny charakter modyfikacji. Rola dziedziczności i środowiska w rozwoju, edukacji i wychowaniu człowieka.
Zmienność modyfikacji odpowiada warunkom siedliskowym i ma charakter adaptacyjny. Takie cechy jak wzrost roślin i zwierząt, ich masa, barwa itp. podlegają zmienności modyfikacji. Pojawienie się zmian modyfikujących wynika z faktu, że warunki środowiskowe wpływają na reakcje enzymatyczne zachodzące w rozwijającym się organizmie iw pewnym stopniu zmieniają ich przebieg.
Ponieważ fenotypowa manifestacja informacji dziedzicznych może być modyfikowana przez warunki środowiskowe, w genotypie organizmu zaprogramowana jest tylko możliwość ich powstania w pewnych granicach, zwanych normą reakcji. Szybkość reakcji reprezentuje granice zmienności modyfikacji cechy dozwolonej dla danego genotypu.
Nasilenie cechy podczas wdrażania genotypu w różnych warunkach nazywa się ekspresywnością. Jest to związane ze zmiennością cechy w normalnym zakresie reakcji.
Ta sama cecha może pojawić się w niektórych organizmach, a nie w innych z tym samym genem. Ilościowy wskaźnik fenotypowej manifestacji genu nazywa się penetracją.
Ekspresję i przenikliwość wspiera dobór naturalny. Podczas badania dziedziczności u ludzi należy pamiętać o obu wzorcach. Zmieniając warunki środowiskowe, można wpływać na penetrację i ekspresję. To, że jeden i ten sam genotyp może być źródłem rozwoju różnych fenotypów, ma zasadnicze znaczenie dla medycyny. Oznacza to, że obciążony nie musi się manifestować. Wiele zależy od warunków, w jakich dana osoba się znajduje. W niektórych przypadkach chorobie jako fenotypowej manifestacji informacji dziedzicznej można zapobiec poprzez przestrzeganie diety lub przyjmowanie leków. Wdrożenie informacji dziedzicznej zależy od środowiska, a powstałe na podstawie historycznie ustalonego genotypu modyfikacje mają zwykle charakter adaptacyjny, ponieważ zawsze są wynikiem reakcji odpowiedzi rozwijający się organizm na czynniki środowiskowe... Charakter zmian mutacyjnych jest inny: są one wynikiem zmian w strukturze cząsteczki DNA, co powoduje zakłócenie ustalonego wcześniej procesu syntezy białek. gdy myszy trzymane są w warunkach podwyższonej temperatury, rodzą potomstwo z wydłużonymi ogonami i powiększonymi uszami. Ta modyfikacja ma charakter adaptacyjny, ponieważ wystające części (ogon i uszy) pełnią w ciele rolę termoregulacyjną: wzrost ich powierzchni umożliwia zwiększenie wymiany ciepła.
Potencjał genetyczny człowieka jest ograniczony w czasie i dość surowy. Jeśli przegapisz termin wczesnej socjalizacji, zniknie, nie mając czasu na realizację. Uderzającym przykładem tego stwierdzenia są liczne przypadki, gdy niemowlęta siłą okoliczności wpadły do dżungli i spędziły kilka lat wśród zwierząt. Po powrocie do społeczności ludzkiej nie mogli już w pełni nadrobić stracony czas: opanowali mowę, nabyli dość złożonych umiejętności działania człowieka, słabo rozwinięte były ich funkcje umysłowe człowieka. Jest to dowód na to, że charakterystyczne cechy ludzkiego zachowania i działalności nabywa się tylko poprzez dziedziczenie społeczne, tylko poprzez przekazanie programu społecznego w procesie edukacji i szkolenia.
Genotypy identyczne (u bliźniąt jednojajowych), znajdujące się w różnych środowiskach, mogą dawać różne fenotypy. Biorąc pod uwagę wszystkie czynniki wpływu, fenotyp człowieka można przedstawić jako składający się z kilku elementów.
Obejmują one: inklinacje biologiczne zakodowane w genach; środowisko (społeczne i przyrodnicze); aktywność jednostki; umysł (świadomość, myślenie).
Interakcja dziedziczności i środowiska w rozwoju człowieka odgrywa ważną rolę przez całe jego życie. Ale nabiera szczególnego znaczenia w okresach formowania się organizmu: embrionalnego, piersiowego, dziecka, dorastania i młodości. W tym czasie zaobserwowano intensywny proces rozwoju organizmu i kształtowania się osobowości.
Dziedziczność określa, czym może się stać organizm, ale człowiek rozwija się pod wpływem obu czynników – zarówno dziedziczności, jak i środowiska. Dziś powszechnie przyjmuje się, że adaptacja człowieka odbywa się pod wpływem dwóch programów dziedziczenia: biologicznego i społecznego. Wszystkie znaki i właściwości każdej osoby są wynikiem interakcji jego genotypu i środowiska. Dlatego każdy człowiek jest zarówno częścią natury, jak i produktem rozwoju społecznego.
91. Zmienność kombinacyjna. Wartość kombinatywnej zmienności w zapewnieniu zróżnicowania genotypowego ludzi: Systemy małżeństwa. Medyczne i genetyczne aspekty rodziny.
Zmienność kombinacyjna związane z uzyskaniem nowych kombinacji genów w genotypie. Osiąga się to w wyniku trzech procesów: a) niezależnej dywergencji chromosomów podczas mejozy; b) ich przypadkowe połączenie podczas zapłodnienia; c) rekombinacja genów dzięki Crossover. Same czynniki dziedziczne (geny) nie zmieniają się, ale pojawiają się nowe ich kombinacje, co prowadzi do pojawienia się organizmów o innych właściwościach genotypowych i fenotypowych. Ze względu na kombinacyjną zmienność u potomstwa powstaje wiele różnych genotypów, które mają: bardzo ważne za proces ewolucyjny ze względu na fakt, że: 1)
różnorodność materiału do procesu ewolucyjnego wzrasta bez zmniejszania żywotności osobników; 2)
rozszerzają się możliwości adaptacji organizmów do zmieniających się warunków środowiskowych, a tym samym zapewniają przetrwanie grupy organizmów (populacji, gatunku) jako całości
Skład i częstość alleli u ludzi w populacjach w dużej mierze zależą od rodzaju małżeństw. W związku z tym duże znaczenie ma badanie typów małżeństw i ich medyczno-genetycznych konsekwencji.
Małżeństwa mogą być: wyborczy, niewybredny.
Aby nie rozróżniać obejmują małżeństwa panmix. Panmixia(grecki nixis - mieszanka) - pół-małżeństwa między osobami o różnych genotypach.
Małżeństwa elektorskie: 1. Krzyżowanie- małżeństwa pomiędzy osobami niepowiązanymi więzami rodzinnymi według ustalonego genotypu, 2.Chów wsobny- małżeństwa pomiędzy krewnymi, 3. Pozytywne-asortymentowe- małżeństwa między osobami o podobnych fenotypach (głuchy i niemy, niewymiarowy z niewyrośniętym, wysoki z wysokim, niedorozwinięty z niedorozwiniętym umysłem itp.). 4. Negatywne-asortymentowe-małżeństwa między osobami o odmiennych fenotypach (głucho-niemy-normalny; niski wysoki; normalny - z piegami itp.). 4 kazirodztwo- małżeństwa między bliskimi krewnymi (między bratem a siostrą).
Małżeństwo wsobne i kazirodztwo jest nielegalne w wielu krajach. Niestety są regiony o wysokiej częstotliwości małżeństw wsobnych. Do niedawna częstość małżeństw inbredowych w niektórych regionach Azji Środkowej sięgała 13-15%.
Znaczenie medyczne i genetyczne małżeństwa wsobne są bardzo negatywne. Przy takich małżeństwach obserwuje się homozygotyzację, częstość chorób autosomalnych recesywnych wzrasta 1,5-2 razy. Populacje wsobne charakteryzują się depresją wsobną, tj. częstotliwość gwałtownie wzrasta, wzrasta częstość niepożądanych alleli recesywnych i wzrasta śmiertelność niemowląt. Małżeństwa pozytywno-asortymentowe również prowadzą do podobnych zjawisk. Krzyżowanie jest genetycznie pozytywne. W przypadku takich małżeństw obserwuje się heterozygotyzację.
92. Zmienność mutacyjna, klasyfikacja mutacji według poziomu zmian w materiale dziedzicznym. Mutacje w komórkach zarodkowych i somatycznych.
Mutacja nazywana jest zmianą spowodowaną reorganizacją struktur reprodukcyjnych, zmianą w aparacie genetycznym. Mutacje występują spazmatycznie i są dziedziczone. W zależności od stopnia zmian w materiale dziedzicznym wszystkie mutacje dzielą się na gen chromosomalny oraz genomowy.
Mutacje genów, lub transgenacje, wpływają na strukturę samego genu. Mutacje mogą zmieniać odcinki cząsteczki DNA o różnej długości. Najmniejsze miejsce, w którym zmiana prowadzi do pojawienia się mutacji, nazywa się mutonem. To może być tylko kilka nukleotydów. Zmiana sekwencji nukleotydów w DNA determinuje zmianę sekwencji trojaczków i ostatecznie program syntezy białek. Należy pamiętać, że naruszenia struktury DNA prowadzą do mutacji tylko wtedy, gdy nie przeprowadza się naprawy.
Mutacje chromosomowe rearanżacje lub aberracje chromosomowe polegają na zmianie liczby lub redystrybucji materiału dziedzicznego chromosomów.
Restrukturyzacja dzieli się na: nutrichromosomalny oraz międzychromosomalny... Rearanżacje wewnątrzchromosomalne polegają na utracie części chromosomu (delecja), podwojeniu lub zwielokrotnieniu niektórych jego odcinków (duplikacja), rotacji fragmentu chromosomu o 180° ze zmianą kolejności ułożenia genów (inwersja).
Mutacje genomowe związane ze zmianą liczby chromosomów. Mutacje genomowe obejmują aneuploidię, haploidię i poliploidię.
Aneuploidia nazywana jest zmiana liczby poszczególnych chromosomów - brak (monosomia) lub obecność dodatkowych (trisomia, tetrasomia, w ogólnym przypadku polisomia) chromosomów, czyli niezrównoważony zestaw chromosomów. Komórki ze zmienioną liczbą chromosomów pojawiają się w wyniku zaburzeń w procesie mitozy lub mejozy, w związku z czym rozróżnia się aneuplodię mitotyczną i mejotyczną. Nazywa się wielokrotnym spadkiem liczby zestawów chromosomów komórek somatycznych w porównaniu z diploidalnym haploidia... Wielokrotny wzrost liczby zestawów chromosomów komórek somatycznych w porównaniu z diploidalnym nazywa się poliploidalność.
Wymienione typy mutacje znajdują się zarówno w komórkach rozrodczych, jak i komórkach somatycznych. Mutacje zachodzące w komórkach zarodkowych nazywane są generatywny... Są przekazywane kolejnym pokoleniom.
Mutacje, które pojawiają się w komórkach ciała na tym czy innym etapie indywidualnego rozwoju organizmu, nazywane są somatyczny... Takie mutacje są dziedziczone tylko przez potomków komórki, w której wystąpiły.
93. Mutacje genów, molekularne mechanizmy występowania, częstość mutacji w przyrodzie. Biologiczne mechanizmy antymutacyjne.
Współczesna genetyka to podkreśla mutacje genów polegają na zmianie struktury chemicznej genów. W szczególności mutacje genów to substytucje, insercje, spadki i utraty par zasad. Najmniejsza część cząsteczki DNA, której zmiana prowadzi do mutacji, nazywana jest mutonem. Jest równy jednej parze nukleotydów.
Istnieje kilka klasyfikacji mutacji genów ... Spontaniczny(spontaniczna) to mutacja, która występuje poza bezpośrednim połączeniem z jakimkolwiek fizycznym lub chemicznym czynnikiem w środowisku.
Jeśli mutacje są powodowane celowo, poprzez narażenie organizmu na czynniki o znanej naturze, nazywa się je wywołany... Czynnik wywołujący mutacje nazywa się mutagen.
Charakter mutagenów jest zróżnicowany są czynnikami fizycznymi, związki chemiczne... Działanie mutagenne niektórych obiektów biologicznych - wirusów, pierwotniaków, robaków pasożytniczych - zostało ustalone, gdy dostają się do organizmu człowieka.
W wyniku mutacji dominujących i recesywnych w fenotypie pojawiają się cechy dominujące i zmienione recesywnie. Dominujący mutacje pojawiają się w fenotypie już w pierwszym pokoleniu. Recesywny mutacje są ukryte w heterozygotach przed działaniem naturalna selekcja w związku z tym gromadzą się w pulach genowych gatunków w dużych ilościach.
Wskaźnikiem intensywności procesu mutacji jest częstość mutacji, która jest obliczana średnio na genom lub osobno dla poszczególnych loci. Średnia częstotliwość mutacji jest porównywalna w wielu organizmach żywych (od bakterii po ludzi) i nie zależy od poziomu i rodzaju organizacji morfofizjologicznej. Jest to równe 10 -4 - 10 -6 mutacji na 1 locus na pokolenie.
Mechanizmy antymutacyjne.
Parowanie chromosomów w diploidalnym kariotypie eukariotycznych komórek somatycznych służy jako czynnik obrony przed niekorzystnymi skutkami mutacji genów. Sparowane geny alleli zapobiegają fenotypowej manifestacji mutacji, jeśli mają one charakter recesywny.
Zjawisko ekstrakopiowania genów kodujących ważne makrocząsteczki przyczynia się do ograniczenia szkodliwych skutków mutacji genów. Na przykład geny rRNA, tRNA, białka histonowe, bez których żywotna aktywność jakiejkolwiek komórki jest niemożliwa.
Wymienione mechanizmy przyczyniają się do zachowania genów wyselekcjonowanych podczas ewolucji, a jednocześnie akumulacji alleli w puli genowej populacji, tworząc rezerwę zmienności dziedzicznej.
94. Mutacje genomowe: poliploidia, haploidia, heteroploidalność. Mechanizmy ich występowania.
Mutacje genomowe są związane ze zmianami liczby chromosomów. Mutacje genomowe obejmują heteroploidalność, haploidia oraz poliploidia.
Poliploidalność- wzrost diploidalnej liczby chromosomów poprzez dodanie całych zestawów chromosomów w wyniku naruszenia mejozy.
W postaciach poliploidalnych następuje wzrost liczby chromosomów, który jest wielokrotnością zestawu haploidalnego: 3n - triploid; 4n - tetraploidalny, 5n - pentaploidalny itp.
Formy poliploidalne różnią się fenotypowo od diploidalnych: wraz ze zmianą liczby chromosomów zmieniają się również właściwości dziedziczne. W poliploidach komórki są zwykle duże; czasami rośliny są gigantyczne.
Formy powstałe w wyniku namnażania się chromosomów jednego genomu nazywane są autoploidami. Znana jest jednak również inna forma poliploidii - alloploidia, w której zwielokrotnia się liczbę chromosomów dwóch różnych genomów.
Nazywa się wielokrotnym spadkiem liczby zestawów chromosomów komórek somatycznych w porównaniu z diploidalnym haploidia... Organizmy haploidalne w naturalnym środowisku spotykane są głównie wśród roślin, w tym wyższych (ropa, pszenica, kukurydza). Komórki takich organizmów mają po jednym chromosomie z każdej pary homologicznej, więc wszystkie allele recesywne pojawiają się w fenotypie. To wyjaśnia zmniejszoną żywotność haploidów.
Heteroploidalność... W wyniku naruszenia mitozy i mejozy liczba chromosomów może się zmienić i nie stać się wielokrotnością zestawu haploidów. Zjawisko, kiedy któryś z chromosomów, zamiast być sparowany, okazuje się być w potrójnej liczbie, otrzymało nazwę trisomie... Jeśli na jednym chromosomie obserwuje się trisomię, wówczas taki organizm nazywa się trisomią, a jego zestaw chromosomów to 2n + 1. Trisomia może znajdować się na dowolnym chromosomie, a nawet na kilku. W przypadku podwójnej trisomii ma zestaw chromosomów 2n + 2, potrójny - 2n + 3 itd.
Odwrotne zjawisko trisomie, tj. nazywana jest utrata jednego z chromosomów z pary w zestawie diploidalnym monosomia, organizm jest monosomiczny; jego wzór genotypowy to 2n-1. W przypadku braku dwóch różnych chromosomów organizm jest podwójnym monosomem o wzorze genotypowym 2n-2 itd.
Z tego, co zostało powiedziane, jasno wynika, że aneuploidia, tj. naruszenie normalnej liczby chromosomów prowadzi do zmian w strukturze i zmniejszenia żywotności organizmu. Im większe naruszenie, tym niższa rentowność. U ludzi naruszenie zrównoważonego zestawu chromosomów prowadzi do bolesnych stanów określanych zbiorczo jako choroby chromosomowe.
Mechanizm występowania mutacje genomowe są związane z patologią naruszenia normalnego rozdziału chromosomów w mejozie, w wyniku czego powstają nieprawidłowe gamety, co prowadzi do mutacji. Zmiany w organizmie są związane z obecnością komórek odmiennych genetycznie.
95. Metody badania ludzkiej dziedziczności. Metody genealogiczne i bliźniacze, ich znaczenie dla medycyny.
Główne metody badania ludzkiej dziedziczności to genealogiczny, bliźniak, populacja-statystyka, metoda dermatoglificzna, cytogenetyczna, biochemiczna, somatyczna genetyka komórek, metoda modelowania
Metoda genealogiczna. Metoda ta opiera się na kompilacji i analizie rodowodów. Rodowód to diagram, który odzwierciedla więzi między członkami rodziny. Analizując rodowody, badają każdy normalny lub (częściej) patologiczny objaw w pokoleniach osób pozostających w więzach rodzinnych.
Metody genealogiczne służą do określenia dziedzicznej lub niedziedzicznej natury cechy, dominacji lub recesywności, mapowania chromosomów, powiązania płci oraz badania procesu mutacji. Z reguły metoda genealogiczna stanowi podstawę wniosków w poradnictwie medycznym genetycznym.
Podczas kompilowania rodowodów stosuje się standardowe oznaczenia. Osoba rozpoczynająca badania jest probantem. Potomek małżeństwa to rodzeństwo, rodzeństwo to rodzeństwo, kuzyni to rodzeństwo kuzynów itp. Potomkowie, którzy mają wspólną matkę (ale różnych ojców) nazywani są samotną macicą, a potomkowie, którzy mają wspólnego ojca (ale różne matki) nazywani są spokrewnionymi; jeśli w rodzinie są dzieci z różnych małżeństw, a ponadto nie mają one wspólnych przodków (np. dziecko z pierwszego małżeństwa matki i dziecko z pierwszego małżeństwa ojca), to nazywa się je połowicznym.
Za pomocą metody genealogicznej można ustalić dziedziczną warunkowość badanej cechy, a także rodzaj jej dziedziczenia. Analizując rodowody na kilku podstawach, można ujawnić powiązany charakter ich dziedziczenia, co jest wykorzystywane przy opracowywaniu map chromosomów. Metoda ta pozwala na badanie intensywności procesu mutacji, ocenę ekspresji i penetracji allelu.
Metoda bliźniacza... Polega na badaniu wzorców dziedziczenia cech u par bliźniąt pojedynczych i podwójnych. Bliźnięta to dwoje lub więcej dzieci, poczętych i urodzonych przez tę samą matkę niemal jednocześnie. Rozróżnij bliźnięta jednojajowe i dwujajowe.
Identyczne (monozygotyczne, identyczne) bliźnięta powstają na najwcześniejszych etapach rozszczepienia zygoty, kiedy dwa lub cztery blastomery zachowują zdolność do rozwinięcia się w pełnoprawny organizm podczas separacji. Ponieważ zygota dzieli się przez mitozę, genotypy bliźniąt jednojajowych są, przynajmniej początkowo, całkowicie identyczne. Bliźnięta jednojajowe są zawsze tej samej płci, w okresie rozwoju wewnątrzmacicznego mają jedno łożysko.
Różne jaja (dizygotyczne, nieidentyczne) pojawiają się, gdy zapłodnione są dwa lub więcej jednocześnie dojrzałych jaj. W ten sposób mają około 50% wspólnych genów. Innymi słowy, są podobni do zwykłych braci i sióstr w swojej budowie genetycznej i mogą być tej samej płci lub płci przeciwnej.
Porównując bliźnięta jednojajowe i dwujajowe wychowywane w tym samym środowisku, można wyciągnąć wniosek o roli genów w rozwoju cech.
Metoda bliźniacza pozwala na wyciąganie świadomych wniosków na temat odziedziczalności cech: roli dziedziczności, środowiska i czynników losowych w określaniu pewnych cech osoby
Profilaktyka i diagnostyka patologii dziedzicznej
Obecnie profilaktyka patologii dziedzicznej odbywa się na czterech poziomach: 1) pregametyczny; 2) prezygotyczny; 3) prenatalne; 4) noworodkowy.
1.) Poziom pregametyczny
Przeprowadzone:
1. Sanitarna kontrola produkcji – wykluczenie wpływu mutagenów na organizm.
2. Zwolnienie kobiet w wieku rozrodczym z pracy przy pracach niebezpiecznych.
3.Tworzenie list choroby dziedziczne które są powszechne w niektórych
terytorium z pok. częsty.
2. Poziom presygotyczny
Najważniejszym elementem tego poziomu profilaktyki jest medyczne poradnictwo genetyczne (MGC) populacji, które informuje rodzinę o stopniu ewentualnego ryzyka urodzenia dziecka z patologią badawczą oraz zapewnia pomoc w podjęciu właściwej decyzji o porodzie.
Poziom prenatalny
Polega na przeprowadzeniu diagnostyki prenatalnej (prenatalnej).
Diagnoza prenatalna To zestaw środków przeprowadzanych w celu określenia dziedzicznej patologii u płodu i przerwania ciąży. Metody diagnostyki prenatalnej obejmują:
1. Skanowanie ultradźwiękowe (USS).
2. Fetoskopia- metoda wizualnej obserwacji płodu w jamie macicy za pomocą elastycznej sondy wyposażonej w układ optyczny.
3... Biopsja kosmówki... Metoda polega na pobraniu kosmków kosmówkowych, hodowli komórek i badaniu ich metodami cytogenetycznymi, biochemicznymi i genetyką molekularną.
4. Amniocenteza- nakłucie płynu owodniowego przez ścianę jamy brzusznej i pobranie
płyn owodniowy. Zawiera komórki płodowe, które można zbadać
cytogenetycznie lub biochemicznie, w zależności od domniemanej patologii płodu.
5. Kordocenteza- nakłucie naczyń pępowiny i pobranie krwi płodowej. Limfocyty płodowe
uprawiane i testowane.
4. Poziom noworodkowy
Na czwartym poziomie noworodki są badane pod kątem wykrycia autosomalnych recesywnych chorób metabolicznych na etapie przedklinicznym, kiedy rozpoczęte w odpowiednim czasie leczenie umożliwia zapewnienie prawidłowego stanu psychicznego i rozwój fizyczny dzieci.
Zasady leczenia chorób dziedzicznych
Istnieją następujące rodzaje leczenia.
1. Objawowy(wpływ na objawy choroby).
2. Patogenetyczne(wpływ na mechanizmy rozwoju choroby).
Leczenie objawowe i patogenetyczne nie eliminuje przyczyn choroby, ponieważ nie eliminuje
wada genetyczna.
W leczeniu objawowym i patogenetycznym można zastosować następujące techniki.
· Korekta wady rozwojowe metodami chirurgicznymi (syndaktyl, polidaktyl,
niezamykanie górnej wargi...
Terapia substytucyjna, której znaczeniem jest wprowadzenie do organizmu
brakujące lub niewystarczające substraty biochemiczne.
· Indukcja metabolizmu- wprowadzenie do organizmu substancji wzmacniających syntezę
niektóre enzymy, a zatem przyspieszają procesy.
· Hamowanie metabolizmu- wprowadzenie do organizmu leków wiążących i usuwających
nieprawidłowe produkty przemiany materii.
· Terapia dietetyczna (żywienie medyczne) – eliminacja z diety substancji, które
nie może być wchłonięty przez organizm.
Perspektywy: W niedalekiej przyszłości genetyka będzie się szybko rozwijać, chociaż nadal jest to w naszych czasach.
bardzo rozpowszechniony w uprawach (hodowla, klonowanie),
medycyna (genetyka medyczna, genetyka mikroorganizmów). Naukowcy mają nadzieję, że w przyszłości
wykorzystaj genetykę do eliminacji wadliwych genów i zwalczania chorób przenoszonych przez
przez dziedziczenie, aby móc leczyć tak poważne choroby jak nowotwory, wirusy
infekcje.
Przy wszystkich niedociągnięciach współczesnej oceny efektu radiogenetycznego nie ma wątpliwości co do powagi konsekwencji genetycznych, jakie czekają ludzkość w przypadku niekontrolowanego wzrostu tła promieniotwórczego w środowisku. Niebezpieczeństwo dalszych testów broni atomowej i wodorowej jest oczywiste.
Jednocześnie wykorzystanie energii atomowej w genetyce i hodowli umożliwia tworzenie nowych metod kontroli dziedziczności roślin, zwierząt i mikroorganizmów, aby lepiej zrozumieć procesy genetycznej adaptacji organizmów. W związku z lotami ludzi w przestrzeń konieczne staje się zbadanie wpływu reakcji kosmicznej na organizmy żywe.
98. Cytogenetyczna metoda diagnozowania anomalii chromosomowych u człowieka. Amniocenteza. Kariotyp i idiogram ludzkich chromosomów. Metoda biochemiczna.
Metoda cytogenetyczna polega na badaniu chromosomów pod mikroskopem. Częściej przedmiotem badań są chromosomy mitotyczne (metafaza), rzadziej mejotyczne (profaza i metafaza). Przy badaniu kariotypów poszczególnych osobników stosuje się metody cytogenetyczne
Przeprowadza się pozyskiwanie materiału rozwijającego się organizmu wewnątrzmacicznego różne sposoby... Jeden z nich jest amniocenteza, za pomocą którego w 15-16 tygodniu ciąży uzyskuje się płyn owodniowy zawierający produkty przemiany materii płodu oraz komórki jego skóry i błon śluzowych
Materiał pobrany podczas amniopunkcji służy do badań biochemicznych, cytogenetycznych i molekularno-chemicznych. Metody cytogenetyczne określają płeć płodu oraz identyfikują mutacje chromosomowe i genomowe. Badanie płynu owodniowego i komórek płodowych metodami biochemicznymi umożliwia wykrycie defektu produktów białkowych genów, ale nie pozwala na określenie lokalizacji mutacji w strukturalnej lub regulacyjnej części genomu. Zastosowanie sond DNA odgrywa ważną rolę w wykrywaniu chorób dziedzicznych i dokładnej lokalizacji uszkodzeń materiału dziedzicznego płodu.
Obecnie za pomocą amniopunkcji diagnozuje się wszystkie nieprawidłowości chromosomalne, ponad 60 dziedzicznych chorób metabolicznych, niezgodność matki i płodu z antygenami erytrocytów.
Nazywa się diploidalny zestaw chromosomów komórki, charakteryzujący się ich liczbą, rozmiarem i kształtem kariotyp... Normalny kariotyp człowieka obejmuje 46 chromosomów, czyli 23 pary: z czego 22 to pary autosomów, a jedna para to chromosomy płci
Aby ułatwić zrozumienie złożonego kompleksu chromosomów tworzących kariotyp, ułożone są one w formie idiogramy... V idiogram chromosomy są ułożone parami w porządku malejącym, wyjątek stanowią chromosomy płci. Największej parze przypisano nr 1, najmniejszej - nr 22. Identyfikacja chromosomów tylko według rozmiaru napotyka ogromne trudności: wiele chromosomów ma podobne rozmiary. Jednak w Ostatnio Dzięki zastosowaniu różnych rodzajów barwników ustalono wyraźne zróżnicowanie ludzkich chromosomów wzdłuż ich długości na paski barwione specjalnymi metodami, a nie barwione. Zdolność do dokładnego różnicowania chromosomów ma ogromne znaczenie dla genetyki medycznej, ponieważ pozwala dokładnie określić charakter naruszeń kariotypu danej osoby.
Metoda biochemiczna
99. Kariotyp i idiogram człowieka. Charakterystyka ludzkiego kariotypu jest normalna
i patologia.
Kariotyp- zestaw znaków (liczba, wielkość, kształt itp.) kompletnego zestawu chromosomów,
tkwiące w komórkach danego gatunku biologicznego (kariotyp gatunkowy), danego organizmu
(kariotyp indywidualny) lub linia (klon) komórek.
Aby określić kariotyp, stosuje się mikrofotografię lub szkic chromosomów z mikroskopią dzielących się komórek.
Każda osoba ma 46 chromosomów, z których dwa to płeć. Kobieta ma dwa chromosomy X
(kariotyp: 46, XX), podczas gdy samce mają jeden chromosom X, a drugi Y (kariotyp: 46, XY). Badanie
Kariotyp wykonuje się techniką zwaną cytogenetyką.
Idiogram- schematyczne przedstawienie haploidalnego zestawu chromosomów organizmu, które
ułożone w rzędzie według ich wielkości, parami w kolejności malejącej. Wyjątek stanowią chromosomy płci, które szczególnie się wyróżniają.
Przykłady najczęstszych nieprawidłowości chromosomalnych.
Zespół Downa to trisomia na 21 parze chromosomów.
Zespół Edwardsa to trisomia na 18 parze chromosomów.
Zespół Patau to trisomia na 13. parze chromosomów.
Zespół Klinefeltera to polisomia chromosomu X u chłopców.
100. Znaczenie genetyki dla medycyny. Cytogenetyczne, biochemiczne, populacyjno-statystyczne metody badania dziedziczności człowieka.
Rola genetyki w życiu człowieka jest bardzo ważna. Jest realizowany przy pomocy medycznego poradnictwa genetycznego. Medyczne poradnictwo genetyczne ma na celu uratowanie ludzkości przed cierpieniem związanym z chorobami dziedzicznymi (genetycznymi). Główne cele medycznego poradnictwa genetycznego to ustalenie roli genotypu w rozwoju danej choroby oraz przewidywanie ryzyka posiadania chorego potomstwa. Zalecenia udzielane w konsultacjach medyczno-genetycznych dotyczących małżeństwa czy prognozy przydatności genetycznej potomstwa mają na celu zapewnienie ich uwzględnienia przez konsultowane osoby, które dobrowolnie podejmują właściwą decyzję.
Metoda cytogenetyczna (kariotypowa). Metoda cytogenetyczna polega na badaniu chromosomów pod mikroskopem. Częściej przedmiotem badań są chromosomy mitotyczne (metafaza), rzadziej mejotyczne (profaza i metafaza). Ta metoda służy również do badania chromatyny płciowej ( cielę barra) Przy badaniu kariotypów poszczególnych osobników stosuje się metody cytogenetyczne
Zastosowanie metody cytogenetycznej pozwala nie tylko na badanie prawidłowej morfologii chromosomów i ogólnie kariotypu, określenie płci genetycznej organizmu, ale przede wszystkim na diagnozowanie różnych chorób chromosomowych związanych ze zmianą liczby chromosomów lub naruszenie ich struktury. Ponadto metoda ta pozwala na badanie procesów mutagenezy na poziomie chromosomów i kariotypu. Jego zastosowanie w poradnictwie medycznym i genetycznym do celów prenatalnej diagnozy chorób chromosomowych umożliwia, poprzez terminowe przerwanie ciąży, zapobieganie pojawieniu się potomstwa z poważnymi zaburzeniami rozwojowymi.
Metoda biochemiczna polega na określeniu we krwi lub moczu aktywności enzymów lub zawartości niektórych produktów przemiany materii. Przez Ta metoda ujawniają zaburzenia metaboliczne ze względu na obecność niekorzystnej kombinacji genów allelicznych w genotypie, częściej alleli recesywnych w stanie homozygotycznym. Dzięki terminowej diagnozie takich chorób dziedzicznych środki zapobiegawcze pozwalają uniknąć poważnych zaburzeń rozwojowych.
Metoda statystyczna populacji. Metoda ta pozwala oszacować prawdopodobieństwo urodzenia osób o określonym fenotypie w danej grupie ludności lub w blisko spokrewnionych małżeństwach; obliczyć częstość nosicielstwa w heterozygotycznym stanie alleli recesywnych. Metoda opiera się na prawie Hardy'ego - Weinberga. Prawo Hardy'ego-Weinberga Jest prawem genetyki populacyjnej. Prawo mówi: „W idealnej populacji częstotliwość genów i genotypów pozostaje stała z pokolenia na pokolenie”.
Główne cechy populacji ludzkich to: wspólne terytorium i możliwość wolnego małżeństwa. Czynniki izolacji, czyli ograniczające swobodę wyboru małżonków, mogą mieć nie tylko bariery geograficzne, ale także religijne i społeczne.
Ponadto metoda ta umożliwia badanie procesu mutacji, roli dziedziczności i środowiska w powstawaniu polimorfizmu fenotypowego u osoby o normalnych cechach, a także w występowaniu chorób, zwłaszcza z dziedziczną predyspozycją. Metodę statystyczną populacji wykorzystuje się do określenia znaczenia czynników genetycznych w antropogenezie, w szczególności w tworzeniu ras.
101. Aberracje strukturalne (aberracje) chromosomów. Klasyfikacja na podstawie zmian w materiale genetycznym. Znaczenie dla biologii i medycyny.
Aberracje chromosomowe powstają w wyniku przegrupowania chromosomów. Są konsekwencją pęknięcia chromosomu, co prowadzi do powstania fragmentów, które następnie łączą się ponownie, ale normalna struktura chromosomu nie zostaje przywrócona. Istnieją 4 główne typy aberracji chromosomowych: braki, podwojenie, inwersja, translokacje, usunięcie- utrata określonego obszaru przez chromosom, który zwykle ulega zniszczeniu
Niedobory powstają z powodu utraty chromosomu w określonym miejscu. Niedobory w środkowej części chromosomu nazywane są delecjami. Utrata znacznej części chromosomu prowadzi organizm do śmierci, utrata nieistotnych obszarów powoduje zmianę właściwości dziedzicznych. Więc. Gdy w kukurydzy brakuje jednego z chromosomów, jej sadzonki są pozbawione chlorofilu.
Podwojenie związane z włączeniem dodatkowej, zduplikowanej części chromosomu. Prowadzi to również do pojawienia się nowych znaków. Tak więc u Drosophila gen oczu z paskami jest spowodowany duplikacją odcinka jednego z chromosomów.
Inwersje obserwuje się, gdy chromosom jest uszkodzony, a odłączony obszar jest obrócony o 180 stopni. Jeśli pęknięcie występuje w jednym miejscu, oderwany fragment jest przyczepiony do chromosomu przeciwnym końcem, jeśli w dwóch miejscach, to środkowy fragment, odwrócony, jest przyczepiony do miejsc pęknięcia, ale z różnymi końcami. Według Darwina inwersje odgrywają ważną rolę w ewolucji gatunków.
Translokacje występują, gdy odcinek chromosomu z jednej pary jest przyłączony do chromosomu niehomologicznego, tj. chromosom z innej pary. Translokacja sekcje jednego z chromosomów są znane u ludzi; może być przyczyną choroby Downa. Większość translokacji obejmujących duże odcinki chromosomów czyni organizm niezdolnym do życia.
Mutacje chromosomowe zmienić dawkę niektórych genów, spowodować redystrybucję genów pomiędzy grupami sprzężeń, zmienić ich lokalizację w grupie sprzężeń. W ten sposób zakłócają równowagę genów komórek organizmu, w wyniku czego dochodzi do odchyleń w rozwoju somatycznym osobnika. Zazwyczaj zmiany dotyczą wielu układów narządów.
Aberracje chromosomowe mają ogromne znaczenie w medycynie. Na aberracje chromosomowe, występuje opóźnienie w ogólnej sprawności fizycznej i rozwój mentalny... Choroby chromosomowe charakteryzują się połączeniem wielu wad wrodzonych. Taka wada jest przejawem zespołu Downa, który obserwuje się w przypadku trisomii w małym odcinku długiego ramienia chromosomu 21. Obraz zespołu płaczu rozwija się wraz z utratą odcinka krótkiego ramienia chromosomu 5. U ludzi najczęściej obserwuje się wady rozwojowe mózgu, układu mięśniowo-szkieletowego, sercowo-naczyniowego i moczowo-płciowego.
102. Pojęcie gatunku, współczesne poglądy na specjację. Zobacz kryteria.
Pogląd Jest zbiorem osobników, które są podobne pod względem kryteriów gatunkowych do tego stopnia, że mogą
naturalnie krzyżują się i wydają płodne potomstwo.
Płodne potomstwo- to, co samo może się rozmnażać. Przykładem bezpłodnego potomstwa jest muł (krzyżówka osła i konia), jest bezpłodny.
Zobacz kryteria- są to znaki, za pomocą których porównuje się 2 organizmy w celu określenia, czy należą one do tego samego gatunku, czy do różnych.
· Morfologiczny – budowa wewnętrzna i zewnętrzna.
· Fizjologiczne i biochemiczne – jak działają narządy i komórki.
· Behawioralne - zachowanie, zwłaszcza w czasie reprodukcji.
Środowisko - zbiór czynników środowiskowych niezbędnych do życia
gatunki (temperatura, wilgotność, żywność, konkurenci itp.)
Geographic – obszar (obszar dystrybucji), tj. terytorium, na którym żyje ten gatunek.
· Genetyczno-rozrodczy - ta sama liczba i struktura chromosomów, która pozwala organizmom wydać płodne potomstwo.
Kryteria widoku są względne, tj. nie można użyć jednego kryterium do oceny gatunku. Na przykład istnieją gatunki rodzeństwa (u komara malarii, u szczurów itp.). Nie różnią się od siebie morfologicznie, ale mają różną liczbę chromosomów i dlatego nie dają potomstwa.
103. Ludność. Jego cechy ekologiczne i genetyczne oraz rola w specjacji.
Populacja- minimalna samoreprodukująca się grupa osobników jednego gatunku, mniej lub bardziej odizolowanych od innych podobnych grup, zamieszkujących określony obszar przez długi szereg pokoleń, tworzących własny system genetyczny i tworzących własną niszę ekologiczną.
Wskaźniki ekologiczne populacji.
Liczba- całkowita liczba osobników w populacji. Wartość ta charakteryzuje się dużym zakresem zmienności, ale nie może być niższa od pewnych granic.
Gęstość- liczba osobników na jednostkę powierzchni lub objętości. Wraz ze wzrostem liczby z reguły rośnie gęstość zaludnienia
Struktura przestrzenna ludność charakteryzuje się osobliwością rozmieszczenia osobników na okupowanym terytorium. Decydują o tym właściwości siedliska i cechy biologiczne gatunku.
Struktura płci odzwierciedla pewien stosunek mężczyzn i kobiet w populacji.
Struktura wiekowa odzwierciedla stosunek różnych grup wiekowych w populacjach, w zależności od oczekiwanej długości życia, czasu dojrzewania, liczby potomstwa.
Wskaźniki genetyczne populacji... Genetycznie populacja charakteryzuje się pulą genów. Jest reprezentowany przez zbiór alleli tworzących genotypy organizmów w danej populacji.
Opisując populacje lub porównując je ze sobą, wykorzystuje się szereg cech genetycznych. Wielopostaciowość... Populację nazywa się polimorficzną w danym locus, jeśli znajdują się w niej dwa lub więcej alleli. Jeśli locus jest reprezentowany przez pojedynczy allel, mówi się o monomorfizmie. Badając wiele loci można określić wśród nich proporcję loci polimorficznych, tj. ocenić stopień polimorfizmu, który jest wskaźnikiem zróżnicowania genetycznego populacji.
Heterozygotyczność... Ważną cechą genetyczną populacji jest heterozygotyczność - częstość występowania heterozygotycznych osobników w populacji. Odzwierciedla również różnorodność genetyczną.
Współczynnik inbredu... Współczynnik ten służy do oszacowania częstości występowania blisko spokrewnionych krzyżówek w populacji.
Stowarzyszenie genów... Częstotliwości alleli różnych genów mogą być od siebie zależne, co charakteryzuje współczynniki asocjacji.
Odległości genetyczne. Różne populacje różnią się częstością alleli. Aby określić ilościowo te różnice, zaproponowano wskaźniki zwane odległościami genetycznymi.
Populacja- elementarna struktura ewolucyjna. W zasięgu dowolnego gatunku osobniki są rozmieszczone nierównomiernie. Obszary gęstego skupienia osobników przeplatają się z przestrzeniami, w których jest ich niewiele lub nie ma ich wcale. W rezultacie powstają mniej lub bardziej izolowane populacje, w których losowe swobodne krzyżowanie (panmixia) występuje systematycznie. Krzyżowanie z innymi populacjami jest bardzo rzadkie i nieregularne. Dzięki panmiksii w każdej populacji powstaje charakterystyczna pula genów, która różni się od pozostałych populacji. To populację należy uznać za elementarną jednostkę procesu ewolucyjnego.
Rola populacji jest wielka, ponieważ w jej obrębie zachodzą prawie wszystkie mutacje. Mutacje te są przede wszystkim związane z izolacją populacji i pulą genów, która różni się ze względu na ich wzajemną izolację. Materiałem do ewolucji jest zmienność mutacyjna, która zaczyna się w populacji, a kończy powstaniem gatunku.