Genetik kod o'z ichiga oladi. Oqsil va nuklein kislotalarning biosintezi
Avvalroq biz nukleotidlar Yerda hayotning paydo bo'lishi uchun muhim xususiyatga ega ekanligini ta'kidlagan edik - agar eritmada bitta polinukleotid zanjiri mavjud bo'lsa, ikkinchi (parallel) zanjirning hosil bo'lish jarayoni o'z-o'zidan o'zaro bog'liq bo'lganlarning to'ldiruvchi bog'lanishi asosida sodir bo'ladi. nukleotidlar. Ikkala zanjirdagi nukleotidlarning bir xil soni va ularning kimyoviy aloqasi bunday reaksiyalarni amalga oshirish uchun zaruriy shartdir. Biroq, oqsil sintezi jarayonida, mRNKdan olingan ma'lumotlar oqsilning tuzilishiga kiritilganda, komplementarlik printsipiga rioya qilish haqida gap bo'lishi mumkin emas. Buning sababi shundaki, mRNK va sintez qilingan oqsilda nafaqat monomerlar soni har xil bo'ladi, balki ular o'rtasida strukturaviy o'xshashlik (bir tomondan, nukleotidlar, boshqa tomondan) yo'qligi ham muhimdir. , aminokislotalar). Ko'rinib turibdiki, bu holda ma'lumotni polinukleotiddan polipeptid tuzilmasiga aniq tarjima qilishning yangi printsipini yaratish kerak bo'ladi. Evolyutsiyada bunday tamoyil yaratildi va uning asosiga genetik kod qo'yildi.
Genetik kod - bu DNK yoki RNKdagi nukleotidlar ketma-ketliklarining ma'lum bir almashinishiga asoslangan nuklein kislota molekulalarida irsiy ma'lumotni qayd etish tizimi bo'lib, ular oqsildagi aminokislotalarga mos keladigan kodonlarni hosil qiladi.
Genetik kod bir nechta xususiyatlarga ega.
Uchlik.
Degeneratsiya yoki ortiqchalik.
Aniqlik.
Polarlik.
Bir-biriga mos kelmaslik.
Kompaktlik.
Ko'p qirralilik.
Shuni ta'kidlash kerakki, ba'zi mualliflar kodga kiritilgan nukleotidlarning kimyoviy xususiyatlari yoki organizm oqsillarida individual aminokislotalarning paydo bo'lish chastotasi va boshqalar bilan bog'liq bo'lgan kodning boshqa xususiyatlarini ham taklif qilishadi. Biroq, bu xususiyatlar yuqoridagilardan kelib chiqadi, shuning uchun biz ularni o'sha erda ko'rib chiqamiz.
a. Uchlik. Genetik kod, ko'plab murakkab tashkil etilgan tizimlar kabi, eng kichik strukturaviy va eng kichik funktsional birlikka ega. Triplet genetik kodning eng kichik tarkibiy birligidir. U uchta nukleotiddan iborat. Kodon genetik kodning eng kichik funktsional birligidir. Qoida tariqasida, mRNK tripletlari kodonlar deb ataladi. Genetik kodda kodon bir nechta funktsiyalarga ega. Birinchidan, uning asosiy vazifasi bitta aminokislotani kodlashidir. Ikkinchidan, kodon aminokislotalarni kodlamasligi mumkin, ammo bu holda u boshqa funktsiyani bajaradi (pastga qarang). Ta'rifdan ko'rinib turibdiki, triplet xarakterlovchi tushunchadir boshlang'ich strukturaviy birlik genetik kod (uch nukleotid). Kodon - xarakterlaydi elementar semantik birlik genom - uchta nukleotid bitta aminokislotaning polipeptid zanjiriga bog'lanishini aniqlaydi.
Elementar strukturaviy birlik dastlab nazariy jihatdan shifrlangan, keyin esa uning mavjudligi eksperimental tarzda tasdiqlangan. Haqiqatan ham, 20 ta aminokislotani bir yoki ikkita nukleotid bilan kodlash mumkin emas. ikkinchisi atigi 4. To'rtta nukleotiddan uchtasi 4 3 = 64 ta variantni beradi, bu tirik organizmlarda mavjud bo'lgan aminokislotalar sonidan ko'proq (1-jadvalga qarang).
64-jadvalda ko'rsatilgan nukleotid birikmalari ikkita xususiyatga ega. Birinchidan, uchliklarning 64 ta variantidan faqat 61 tasi kodon va har qanday aminokislotalarni kodlaydi, ular deyiladi. sezuvchi kodonlar... Uchta uchlik kodlanmaydi
1-jadval.
Messenger RNK kodonlari va tegishli aminokislotalar
|
B o n va I c o d o n o v |
|||||
|
Bema'nilik |
Bema'nilik |
||||
|
Bema'nilik | |||||
|
uchrashdi |
|||||
|
Mil |
|||||
aminokislotalar a translatsiyaning tugashini ko'rsatuvchi to'xtash signallari. Bunday uchta uchlik bor - UAA, UAG, UGA, ular "ma'nosiz" (bema'ni kodonlar) deb ham ataladi. Tripletdagi bir nukleotidni boshqasiga almashtirish bilan bog'liq bo'lgan mutatsiya natijasida sezgi kodonidan ma'nosiz kodon paydo bo'lishi mumkin. Ushbu turdagi mutatsiya deyiladi bema'ni mutatsiya... Agar bunday to'xtash signali gen ichida (uning axborot qismida) hosil bo'lsa, u holda bu joyda protein sintezi paytida jarayon doimiy ravishda to'xtatiladi - oqsilning faqat birinchi (to'xtash signalidan oldin) qismi sintezlanadi. Ushbu patologiyaga ega bo'lgan odamda protein etishmasligi va bu etishmasligi bilan bog'liq alomatlar bo'ladi. Masalan, bunday mutatsiya gemoglobinning beta zanjirini kodlovchi genda topilgan. Qisqartirilgan faol bo'lmagan gemoglobin zanjiri sintezlanadi, u tezda yo'q qilinadi. Natijada, beta zanjiridan mahrum bo'lgan gemoglobin molekulasi hosil bo'ladi. Bunday molekulaning o'z vazifalarini to'liq bajarishi dargumon. Gemolitik anemiya (beta-nol talassemiya, yunoncha "Talas" so'zidan - O'rta er dengizi, bu kasallik birinchi marta kashf etilgan) sifatida rivojlanadigan jiddiy kasallik paydo bo'ladi.
To'xtash kodonlarining ta'sir mexanizmi hissiy kodonlardan farq qiladi. Bu aminokislotalarni kodlovchi barcha kodonlar uchun mos keladigan tRNKlar topilganligidan kelib chiqadi. Bema'ni kodonlar uchun tRNK topilmadi. Binobarin, tRNK oqsil sintezini to'xtatish jarayonida ishtirok etmaydi.
KodonAVG (bakteriyalarda, ba'zan GUG) nafaqat metionin va valin aminokislotalarini kodlaydi, balkieshittirish tashabbuskori .
b. Degeneratsiya yoki ortiqchalik.
64 ta uchlikdan 61 tasi 20 ta aminokislotalarni kodlaydi. Aminokislotalar sonidan uch barobar ko'pligi ma'lumot uzatishda ikkita kodlash variantidan foydalanish mumkinligini ko'rsatadi. Birinchidan, 20 ta aminokislotalarni kodlashda 64 ta kodonning hammasi ham ishtirok eta olmaydi, faqat 20 tasi, ikkinchidan, aminokislotalar bir nechta kodonlar bilan kodlanishi mumkin. Tadqiqotlar shuni ko'rsatdiki, tabiat oxirgi variantdan foydalangan.
Uning afzalligi aniq. Agar aminokislotalarni kodlashda tripletlarning 64 ta variantidan faqat 20 tasi ishtirok etgan bo'lsa, u holda 44 ta uchlik (64 tadan) kodlanmagan bo'lib qoladi, ya'ni. ma'nosiz (bema'ni kodonlar). Avvalroq, mutatsiya natijasida kodlovchi tripletning bema'ni kodonga aylanishi hujayra hayoti uchun qanchalik xavfli ekanligini ta'kidlagan edik - bu RNK polimerazasining normal ishlashini sezilarli darajada buzadi va natijada kasalliklarning rivojlanishiga olib keladi. Hozirgi vaqtda bizning genomimizda uchta kodon ma'nosiz, ammo endi ma'nosiz kodonlar soni taxminan 15 baravar ko'paysa nima bo'lishini tasavvur qiling. Bunday vaziyatda oddiy kodonlardan bema'ni kodonlarga o'tish beqiyos darajada yuqori bo'lishi aniq.
Bitta aminokislota bir nechta tripletlar bilan kodlangan kod degenerativ yoki ortiqcha deb ataladi. Bir nechta kodonlar deyarli har bir aminokislotaga to'g'ri keladi. Shunday qilib, leytsin aminokislotasini oltita uchlik - UUA, UUG, CUU, CUTS, CUA, CUG bilan kodlash mumkin. Valin to'rtta uchlik, fenilalanin ikkita va faqat kodlangan triptofan va metionin bitta kodon bilan kodlangan. Turli xil belgilar bilan bir xil ma'lumotlarni yozib olish bilan bog'liq xususiyat deyiladi degeneratsiya.
Bitta aminokislotaga biriktirilgan kodonlar soni oqsillarda aminokislotalarning paydo bo'lish chastotasi bilan yaxshi bog'liq.
Va bu, ehtimol, tasodifiy emas. Proteinda aminokislotalarning paydo bo'lish chastotasi qanchalik yuqori bo'lsa, genomda ushbu aminokislotalarning kodoni qanchalik tez-tez namoyon bo'lsa, uning mutagen omillar bilan zararlanish ehtimoli shunchalik yuqori bo'ladi. Shu sababli, mutatsiyaga uchragan kodonning yuqori degeneratsiyasi bilan bir xil aminokislotalarni kodlash imkoniyati ko'proq ekanligi aniq. Ushbu pozitsiyalardan genetik kodning degeneratsiyasi inson genomini shikastlanishdan himoya qiluvchi mexanizmdir.
Shuni ta'kidlash kerakki, degeneratsiya atamasi molekulyar genetikada va boshqa ma'noda qo'llaniladi. Shunday qilib, kodondagi ma'lumotlarning asosiy qismi dastlabki ikkita nukleotidga to'g'ri keladi, kodonning uchinchi pozitsiyasidagi asos ahamiyatsiz bo'lib chiqadi. Ushbu hodisa "uchinchi bazaning degeneratsiyasi" deb ataladi. Oxirgi xususiyat mutatsiyalarning ta'sirini kamaytiradi. Masalan, qizil qon hujayralarining asosiy vazifasi kislorodni o'pkadan to'qimalarga va karbonat angidridni to'qimalardan o'pkaga olib borishi ma'lum. Bu funktsiyani nafas olish pigmenti - gemoglobin bajaradi, u eritrotsitning butun sitoplazmasini to'ldiradi. U tegishli gen tomonidan kodlangan protein qismi - globindan iborat. Proteindan tashqari, gemoglobin molekulasi tarkibiga temir o'z ichiga olgan gem kiradi. Globin genlaridagi mutatsiyalar turli xil gemoglobin variantlarining paydo bo'lishiga olib keladi. Ko'pincha mutatsiyalar bilan bog'liq bir nukleotidni boshqasi bilan almashtirish va genda yangi kodon paydo bo'lishi, bu gemoglobin polipeptid zanjirida yangi aminokislotalarni kodlashi mumkin. Tripletda mutatsiya natijasida har qanday nukleotid almashtirilishi mumkin - birinchi, ikkinchi yoki uchinchi. Bir necha yuz mutatsiyalar globin genlarining yaxlitligiga ta'sir qilishi ma'lum. Haqida 400 bulardan gendagi yagona nukleotidlarning almashinishi va polipeptidda mos keladigan aminokislotalarning o'rnini bosishi bilan bog'liq. Bulardan faqat 100 almashtirishlar gemoglobinning beqarorligiga va engildan o'ta og'irgacha bo'lgan turli xil kasalliklarga olib keladi. 300 (taxminan 64%) almashtirish mutatsiyasi gemoglobin funktsiyasiga ta'sir qilmaydi va patologiyaga olib kelmaydi. Buning sabablaridan biri serin, leysin, prolin, arginin va boshqa ba'zi aminokislotalarni kodlovchi tripletda uchinchi nukleotidning almashinishi kodon sinonimining paydo bo'lishiga olib kelganda, yuqorida aytib o'tilgan "uchinchi asosning degeneratsiyasi" dir. bir xil aminokislotalarni kodlash. Fenotipik jihatdan bu mutatsiya paydo bo'lmaydi. Bundan farqli o'laroq, 100% hollarda birinchi yoki ikkinchi nukleotidni tripletda har qanday almashtirish gemoglobinning yangi variantining paydo bo'lishiga olib keladi. Ammo bu holatda ham jiddiy fenotipik buzilishlar bo'lmasligi mumkin. Buning sababi gemoglobindagi aminokislotaning fizik-kimyoviy xossalari bo'yicha birinchisiga o'xshash boshqasi bilan almashtirilishidir. Masalan, agar gidrofil xossaga ega bo'lgan aminokislota o'rniga xuddi shunday xususiyatlarga ega bo'lgan boshqa aminokislota bo'lsa.
Gemoglobin gemning temir porfirin guruhidan (kislorod va karbonat angidrid molekulalari biriktiriladi) va oqsil - globindan iborat. Katta yoshli gemoglobin (HbA) ikkita bir xil gemoglobinni o'z ichiga oladi -zanjirlar va ikkita -zanjirlar. Molekula -zanjirda 141 ta aminokislota qoldiqlari mavjud; -zanjir - 146, - va b-zanjirlar ko'p aminokislotalar qoldiqlarida farqlanadi. Har bir globin zanjirining aminokislotalar ketma-ketligi o'z geni tomonidan kodlangan. Genni kodlash - zanjir 16-xromosomaning qisqa qo'lida joylashgan; -gen - 11-xromosomaning qisqa qo'lida. Genlarni kodlashda almashtirish - birinchi yoki ikkinchi nukleotidning gemoglobin zanjiri deyarli har doim oqsilda yangi aminokislotalarning paydo bo'lishiga, gemoglobinning disfunktsiyasiga va bemor uchun og'ir oqibatlarga olib keladi. Masalan, CAU (gistidin) uchliklaridan birida "C" ni "Y" ga almashtirish boshqa aminokislota - tirozinni kodlaydigan CAU ning yangi tripletining paydo bo'lishiga olib keladi. Gistidin polipeptidining tirozinga bo'lgan b-zanjirlari gemoglobinni beqarorlashtiradi. Kasallik methemoglobinemiyani rivojlantiradi. Mutatsiya natijasida glutamik kislotani 6-pozitsiyadagi valin bilan almashtirish -zanjirlar eng jiddiy kasallikning sababi - o'roqsimon hujayrali anemiya. Keling, qayg'uli ro'yxatni davom ettirmaylik. Biz shuni ta'kidlaymizki, birinchi ikkita nukleotid almashtirilganda, aminokislota avvalgisiga o'xshash fizik-kimyoviy xususiyatlarga ega bo'lishi mumkin. Shunday qilib, glutamik kislotani (GAA) kodlovchi tripletlardan birida 2-nukleotidni almashtirish -"Y" bilan zanjir valinni kodlovchi yangi triplet (GUA) paydo bo'lishiga olib keladi va birinchi nukleotidning "A" bilan almashtirilishi amino kislotalar lizinni kodlaydigan AAA tripletini hosil qiladi. Glutamik kislota va lizin fizik-kimyoviy xossalari bo'yicha o'xshash - ikkalasi ham gidrofildir. Valin hidrofobik aminokislotadir. Shuning uchun gidrofil glutamik kislotani hidrofobik valin bilan almashtirish gemoglobinning xususiyatlarini sezilarli darajada o'zgartiradi, bu oxir-oqibat o'roqsimon hujayrali anemiya rivojlanishiga olib keladi, gidrofil glutamik kislotani gidrofil lizin bilan almashtirish esa gemoglobin funktsiyasini kamroq darajada o'zgartiradi - bemorlar engil shaklga ega. anemiyadan. Uchinchi asosni almashtirish natijasida yangi triplet avvalgisi kabi bir xil aminokislotalarni kodlashi mumkin. Misol uchun, agar CAC tripletida urasil sitozin bilan almashtirilgan bo'lsa va CAC triplet paydo bo'lsa, u holda odamlarda deyarli hech qanday fenotipik o'zgarishlar aniqlanmaydi. Bu tushunarli, chunki ikkala uchlik ham bir xil aminokislota, histidinni kodlaydi.
Xulosa qilib shuni ta'kidlash joizki, genetik kodning degeneratsiyasi va uchinchi bazaning umumiy biologik nuqtai nazardan degeneratsiyasi DNK va RNKning noyob tuzilishida evolyutsiyaga kiritilgan himoya mexanizmlari hisoblanadi.
v. Aniqlik.
Har bir triplet (ma'nosizlardan tashqari) faqat bitta aminokislotalarni kodlaydi. Shunday qilib, kodon - aminokislota yo'nalishi bo'yicha, genetik kod bir ma'noli, aminokislota - kodon yo'nalishi bo'yicha noaniq (degeneratsiya) bo'ladi.
Aniq
Aminokislota kodoni
Degeneratsiya
Va bu holda, genetik kodda noaniqlik zarurati aniq. Boshqa bir variantda, bir xil kodonni tarjima qilish jarayonida oqsil zanjiriga turli xil aminokislotalar kiritiladi va natijada birlamchi tuzilmalari va funktsiyalari turlicha bo'lgan oqsillar hosil bo'ladi. Hujayra metabolizmi "bir gen - bir nechta poipeptidlar" ish rejimiga o'tadi. Bunday vaziyatda genlarning tartibga solish funktsiyasi butunlay yo'qolishi aniq.
Polarlik
DNK va mRNK dan ma'lumotni o'qish faqat bitta yo'nalishda sodir bo'ladi. Qutblilik yuqori tartibli tuzilmalarni (ikkilamchi, uchinchi darajali va boshqalarni) aniqlash uchun zarurdir. Biz avvalroq quyi tartibli tuzilmalar yuqori tartibli tuzilmalarni belgilashini muhokama qilgan edik. Oqsillardagi uchinchi darajali tuzilish va yuqori tartibli tuzilmalar sintezlangan RNK zanjiri DNK molekulasidan yoki polipeptid zanjiri ribosomadan chiqib ketishi bilanoq darhol hosil bo'ladi. RNK yoki polipeptidning erkin uchi uchinchi darajali tuzilishga ega bo'lsa-da, zanjirning ikkinchi uchi hali ham DNKda (agar RNK transkripsiyalangan bo'lsa) yoki ribosomada (agar polipeptid transkripsiyalangan bo'lsa) sintezlanadi.
Shuning uchun ma'lumotni o'qishning bir yo'nalishli jarayoni (RNK va oqsil sintezida) nafaqat sintezlangan moddadagi nukleotidlar yoki aminokislotalarning ketma-ketligini aniqlash uchun, balki ikkilamchi, uchinchi darajali va boshqalarni qattiq aniqlash uchun juda muhimdir. tuzilmalar.
e) Bir-biriga mos kelmaslik.
Kod bir-biriga o'xshash va bir-biriga mos kelmaydigan bo'lishi mumkin. Aksariyat organizmlarda bir-biriga o'xshash kod mavjud emas. Ba'zi faglarda bir-biriga o'xshash kod mavjud.
Bir-birining ustiga chiqmaydigan kodning mohiyati shundan iboratki, bir kodonning nukleotidi bir vaqtning o'zida boshqa kodonning nukleotidi bo'la olmaydi. Agar kod bir-birining ustiga chiqqan bo'lsa, etti nukleotidlar ketma-ketligi (GCCHCUG) ikkita aminokislotani (alanin-alanin) kodlashi mumkin edi (33-rasm, A), bir-biriga mos kelmaydigan koddagi kabi, uchta (agar bitta bo'lsa). nukleotid keng tarqalgan) (33-rasm, B) yoki beshta (agar ikkita nukleotid umumiy bo'lsa) (33-rasm, C ga qarang). Oxirgi ikki holatda har qanday nukleotidning mutatsiyasi ikki, uch va hokazo ketma-ketlikning buzilishiga olib keladi. aminokislotalar.
Shu bilan birga, bitta nukleotid mutatsiyasi har doim bitta aminokislotaning polipeptid tarkibiga kirishini buzishi aniqlandi. Bu bir-biriga mos kelmaydigan kod uchun muhim dalil.
Buni 34-rasmda tushuntirib beraylik. Qalin chiziqlar bir-birining ustiga chiqmagan va bir-birining ustiga chiqayotgan kod holatlarida aminokislotalarni kodlovchi tripletlarni ko'rsatadi. Tajribalar genetik kodning bir-biriga mos kelmasligini aniq ko'rsatdi. Tajriba tafsilotlariga kirmasdan, shuni ta'kidlaymizki, agar biz nukleotidlar ketma-ketligidagi uchinchi nukleotidni almashtirsak (34-rasmga qarang).bor (yulduzcha bilan belgilangan) boshqa narsaga:
1. Bir-biriga mos kelmaydigan kod bilan, ushbu ketma-ketlik bilan boshqariladigan oqsil bitta (birinchi) aminokislota (yulduzcha bilan belgilangan) o'rnini bosadi.
2. A variantida bir-biriga o'xshash kod bilan ikkita (birinchi va ikkinchi) aminokislotalarda (yulduzcha bilan belgilangan) o'zgarishlar yuz beradi. B variantida almashtirish uchta aminokislotaga ta'sir qilgan bo'lardi (yulduzcha bilan belgilangan).
Biroq, ko'plab tajribalar shuni ko'rsatdiki, DNKdagi bitta nukleotid buzilganda, oqsildagi buzilishlar doimo bir-birining ustiga chiqmaydigan kodga xos bo'lgan faqat bitta aminokislotaga taalluqlidir.
GtsUGtsUG GtsUGtsUG GtsUGtsUG
GTsU GTsU GTsU UGTs TsUG GTsU TsUG UGTs GtsU TsUG
*** *** *** *** *** ***
Alanin - Alanin Ala - Cis - Lei Ala - Lei - Lei - Ala - Lei
A B C
Bir-biriga mos kelmaydigan kod Bir-biriga mos keladigan kod
Guruch. 34. Genomda bir-biriga mos kelmaydigan kod mavjudligini tushuntiruvchi sxema (matndagi tushuntirish).
Genetik kodning bir-biriga mos kelmasligi boshqa xususiyat bilan bog'liq - ma'lumotni o'qish ma'lum bir nuqtadan boshlanadi - boshlash signali. mRNKdagi bunday boshlash signali AUG metioninni kodlovchi kodondir.
Shuni ta'kidlash kerakki, odamlarda hali ham umumiy qoidadan chetga chiqadigan va bir-biriga mos keladigan kam sonli genlar mavjud.
e) ixchamlik.
Kodonlar orasida tinish belgilari yo'q. Boshqacha qilib aytganda, tripletlar bir-biridan, masalan, bitta ma'nosiz nukleotid bilan ajratilmaydi. Genetik kodda "tinish belgilari" yo'qligi tajribalarda isbotlangan.
f. Ko'p qirralilik.
Kod Yerda yashovchi barcha organizmlar uchun bir xil. To'g'ridan-to'g'ri dalillar genetik kodning universalligi DNK ketma-ketliklarini mos keladigan oqsil ketma-ketliklari bilan solishtirish orqali olingan. Ma'lum bo'lishicha, bir xil kod qiymatlari to'plami barcha bakterial va eukaryotik genomlarda qo'llaniladi. Istisnolar bor, lekin ko'p emas.
Genetik kodning universalligiga birinchi istisnolar ba'zi hayvonlar turlarining mitoxondriyalarida topilgan. Bu UGA terminatori kodoni bilan bog'liq bo'lib, u aminokislota triptofanni kodlovchi UGG kodoniga o'xshab o'qiladi. Universallikdan boshqa kam uchraydigan og'ishlar topildi.
MH. Genetik kod - DNK yoki RNKdagi nukleotidlar ketma-ketliklarining ma'lum bir almashinishiga asoslangan nuklein kislota molekulalarida irsiy ma'lumotni qayd qilish tizimi;
oqsil tarkibidagi aminokislotalarga mos keladi.Genetik kod bir nechta xususiyatlarga ega.
Tanadagi metabolizmda yetakchi rol
oqsillar va nuklein kislotalarga tegishli.
Protein moddalari barcha hayotiy hujayra tuzilmalarining asosini tashkil qiladi, g'ayrioddiy yuqori reaktivlikka ega va katalitik funktsiyalarga ega.
Nuklein kislotalar hujayraning eng muhim organi - yadroning bir qismi, shuningdek, sitoplazma, ribosomalar, mitoxondriyalar va boshqalar. Nuklein kislotalar organizmning irsiyatida, o'zgaruvchanligida, oqsil sintezida muhim, eng muhim rol o'ynaydi.
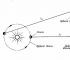
Reja sintez oqsil hujayra yadrosida saqlanadi va sintez bevosita yadrodan tashqarida sodir bo'ladi, shuning uchun zarur yetkazib berish xizmati kodlangan reja yadrodan sintez joyigacha. Ushbu yetkazib berish xizmati RNK molekulalari tomonidan amalga oshiriladi.
Jarayon soatda boshlanadi yadro hujayralar: DNKning bir qismi "narvon" ochiladi va ochiladi. Buning yordamida RNK harflari bilan bog'lanish hosil bo'ladi ochiq xatlar DNK DNK zanjirlaridan biridir. Ferment RNK harflarini bir ipga birlashtirish uchun o'tkazadi. DNK harflari RNK harflariga shunday "qayta yoziladi". Yangi hosil bo'lgan RNK zanjiri ajralib chiqadi va DNK "narvon" yana aylanadi. DNKdan ma'lumotni o'qish va uni RNK matritsasidan sintez qilish jarayoni deyiladi transkripsiya , va sintezlangan RNK informatsion yoki deyiladi i-RNK .
Keyingi o'zgarishlardan so'ng, bunday kodlangan i-RNK tayyor bo'ladi. i-RNK yadrodan chiqadi va oqsil sintezi joyiga boradi, u erda i-RNK harflari dekodlanadi. Har bir uchta harfdan iborat i-RNK ma'lum bir aminokislotani ifodalovchi "harf" ni hosil qiladi.
RNKning yana bir turi bu aminokislotani qidiradi, uni ferment yordamida ushlaydi va oqsil sintezi joyiga yetkazadi. Bu RNK transport RNK yoki t-RNK deb ataladi. i-RNK xabari o'qilishi va tarjima qilinishi bilan aminokislotalar zanjiri o'sadi. Bu zanjir buralib, bir xil oqsil hosil qilish uchun noyob shaklga aylanadi. Hatto oqsillarni katlama jarayoni ham diqqatga sazovordir: hamma narsani kompyuter yordamida hisoblash variantlari 100 ta aminokislotadan tashkil topgan oʻrta kattalikdagi oqsilni buklash uchun 1027 (!) yil kerak boʻladi. Va organizmda 20 ta aminokislotadan iborat zanjir hosil bo'lishi uchun bir soniyadan ko'proq vaqt ketadi va bu jarayon tananing barcha hujayralarida doimiy ravishda sodir bo'ladi.
Genlar, genetik kod va uning xossalari.
Yer yuzida 7 milliardga yaqin odam yashaydi. 25-30 million juft bir xil egizaklardan tashqari, genetik jihatdan hamma odamlar har xil : har biri o'ziga xos, o'ziga xos irsiy xususiyatlarga ega, xarakter xususiyatlari, qobiliyatlari, temperamenti.
Bunday farqlar tushuntiriladi genotiplardagi farqlar- organizm genlari to'plami; har biri o'ziga xosdir. Muayyan organizmning genetik belgilari mujassamlangan oqsillarda - shuning uchun bir odamning oqsilining tuzilishi boshqa odamning oqsilidan biroz farq qiladi.
Bu degani emas odamlarda aynan bir xil oqsillar yo'qligi. Xuddi shu funktsiyalarni bajaradigan oqsillar bir xil bo'lishi mumkin yoki bir-biridan bir yoki ikkita aminokislota bilan ozgina farq qilishi mumkin. Lekin mavjud emas Yerdagi odamlar (bir xil egizaklar bundan mustasno), ularda barcha oqsillar mavjud bir xil .
Oqsilning birlamchi tuzilishi haqida ma'lumot DNK molekulasi hududidagi nukleotidlar ketma-ketligi sifatida kodlangan, gen - organizmning irsiy axborot birligi. Har bir DNK molekulasida ko'plab genlar mavjud. Buni organizmning barcha genlarining yig'indisi tashkil qiladi genotip ... Shunday qilib,
Gen - bu DNKning alohida bo'limiga mos keladigan organizmning irsiy ma'lumotlar birligi
Irsiy ma'lumotlar yordamida kodlangan genetik kod , bu barcha organizmlar uchun universal bo'lib, faqat genlarni hosil qiluvchi nukleotidlarning almashinishida va o'ziga xos organizmlarning oqsillarini kodlashda farqlanadi.
Genetik kod turli ketma-ketlikda (AAT, HCA, ACG, THC va boshqalar) birlashtirilgan DNK nukleotidlarining tripletlari (uchliklari) dan iborat bo'lib, ularning har biri o'ziga xos aminokislotalarni (polipeptid zanjiriga kiritiladi) kodlaydi.
Aslida kod
hisobga oladi i-RNK molekulasidagi nukleotidlar ketma-ketligi
beri u DNKdan ma'lumotni olib tashlaydi (jarayon transkripsiyalar
) va uni sintezlangan oqsillar molekulalaridagi aminokislotalar ketma-ketligiga aylantiradi (jarayon eshittirishlar
).
i-RNK tarkibiga A-C-G-U nukleotidlari kiradi, ularning tripletlari deyiladi. kodonlar
: i-RNKdagi DNK CGT dagi triplet HCA tripletiga, AAG DNK tripleti esa UUC tripletiga aylanadi. Aynan i-RNK kodonlari
genetik kod yozuvda aks ettirilgan.
Shunday qilib, genetik kod - nuklein kislota molekulalarida nukleotidlar ketma-ketligi ko'rinishidagi irsiy ma'lumotlarni qayd qilish uchun yagona tizim. ... Genetik kod azotli asoslarda farq qiluvchi faqat to'rtta harf - nukleotidlardan iborat alifbodan foydalanishga asoslangan: A, T, G, C.
Genetik kodning asosiy xususiyatlari:

1. Genetik kod uchlik... Triplet (kodon) - bitta aminokislotalarni kodlaydigan uchta nukleotidlar ketma-ketligi. Proteinlar 20 ta aminokislotadan iborat bo'lganligi sababli, ularning har birini bitta nukleotid bilan kodlash mumkin emasligi aniq. DNKda faqat to'rt turdagi nukleotidlar mavjud bo'lganligi sababli, bu holda 16 ta aminokislotalar kodlanmagan holda qoladi.). Aminokislotalarni kodlash uchun ikkita nukleotid ham yo'q, chunki bu holda faqat 16 ta aminokislotani kodlash mumkin. Ma'nosi, eng kichik raqam bitta aminokislotani kodlaydigan nukleotidlar kamida uchta bo'lishi kerak. Bunday holda, nukleotidlarning mumkin bo'lgan tripletlari soni 43 = 64 ni tashkil qiladi.
2. Ortiqchalik (degeneratsiya) kod uning triplet tabiatining natijasidir va bitta aminokislota bir nechta tripletlar bilan kodlanishi mumkinligini anglatadi (chunki 20 ta aminokislota va 64 ta triplet mavjud), faqat bitta triplet bilan kodlangan metionin va triptofan bundan mustasno. Bundan tashqari, ba'zi tripletlar o'ziga xos funktsiyalarni bajaradilar: i-RNK molekulasida UAA, UAG, UGA tripletlari tugatish kodonlari, ya'ni. STOP-polipeptid zanjiri sintezini to'xtatuvchi signallar. DNK zanjirining boshida joylashgan metionin (AUG) ga mos keladigan triplet aminokislotalarni kodlamaydi, balki o'qishni boshlash (qo'zg'atish) funktsiyasini bajaradi.
3. Aniqlik kod - ortiqcha bilan bir qatorda kod xususiyatga ega noaniqlik : har bir kodon faqat mos keladi bitta o'ziga xos aminokislota.
4. Kollinearlik kod, ya'ni. gen nukleotidlar ketma-ketligi aynan oqsildagi aminokislotalarning ketma-ketligiga mos keladi.
5. Genetik kod bir-birining ustiga chiqmaydigan va ixcham , ya'ni unda "tinish belgilari" mavjud emas. Bu shuni anglatadiki, o'qish jarayoni ustunlar (uchlik) bir-birining ustiga tushishiga yo'l qo'ymaydi va ma'lum bir kodondan boshlab, o'qish doimiy ravishda uch marta uch martagacha davom etadi. STOP-signallar ( tugatish kodonlari).
6. Genetik kod ko'p tomonli ya'ni barcha organizmlarning yadro genlari bu organizmlarning tashkiliy darajasi va tizimli joylashuvidan qat'i nazar, oqsillar haqidagi ma'lumotlarni bir xil tarzda kodlaydi.
Mavjud genetik kod jadvallari shifrni ochish uchun kodonlar i-RNK va oqsil molekulalarining qurilish zanjirlari.

Matritsa sintez reaksiyalari.
Tirik tizimlarda jonsiz tabiatda noma'lum reaktsiyalar mavjud - matritsa sintezi reaksiyalari.
"matritsa" atamasi texnologiyada ular tangalar, medallar, tipografik turlarni quyish uchun ishlatiladigan shaklni bildiradi: qotib qolgan metall quyish uchun ishlatilgan shaklning barcha tafsilotlarini aniq takrorlaydi. Matritsa sintezi matritsadagi quymaga o'xshaydi: yangi molekulalar allaqachon mavjud bo'lgan molekulalarning tuzilishida belgilangan rejaga qat'iy muvofiq sintezlanadi.
Matritsa printsipi yotadi asosiyda hujayraning eng muhim sintetik reaktsiyalari, masalan, nuklein kislotalar va oqsillar sintezi. Bu reaksiyalar sintez qilingan polimerlardagi monomer birliklarining aniq, qat'iy o'ziga xos ketma-ketligini ta'minlaydi.
Bu erda yo'nalish monomerlarni ma'lum bir joyga tortib olish hujayralar - matritsa bo'lib xizmat qiladigan molekulalarda, bu erda reaksiya davom etadi. Agar bunday reaktsiyalar molekulalarning tasodifiy to'qnashuvi natijasida sodir bo'lsa, ular cheksiz sekinlik bilan boradi. Matritsa printsipiga asoslangan murakkab molekulalarning sintezi tez va aniq. Matritsaning roli nuklein kislota makromolekulalari matritsa reaktsiyalarida o'ynaydi DNK yoki RNK .

Monomerik molekulalar polimer sintez qilinadigan nukleotidlar yoki aminokislotalar - komplementarlik printsipiga muvofiq matritsada qat'iy belgilangan, belgilangan tartibda joylashadi va o'rnatiladi.
Keyin sodir bo'ladi monomer birliklarining polimer zanjiriga "o'zaro bog'lanishi" va tayyor polimer matritsadan tashlanadi.
Bundan keyin matritsa tayyor yangi polimer molekulasining yig'ilishiga. Ma'lumki, ma'lum bir shaklda faqat bitta tanga, bitta harf quyilishi mumkin bo'lganidek, ma'lum bir matritsa molekulasida faqat bitta polimerni "yig'ish" mumkin.
Reaksiyalarning matritsa turi- tirik tizimlar kimyosining o'ziga xos xususiyati. Ular barcha tirik mavjudotlarning asosiy xususiyati - uning o'z turini ko'paytirish qobiliyatining asosidir.
Matritsa sintez reaksiyalari
1. DNK replikatsiyasi - replikatsiya (lotincha replicatio - yangilanish) - asosiy DNK molekulasi matritsasida dezoksiribonuklein kislotaning qiz molekulasini sintez qilish jarayoni. Ona hujayraning keyingi bo'linishi paytida har bir qiz hujayra DNK molekulasining bir nusxasini oladi, bu asl ona hujayraning DNKsi bilan bir xil bo'ladi. Bu jarayon irsiy axborotning nasldan naslga toʻgʻri uzatilishini taʼminlaydi. DNK replikatsiyasi 15-20 xil oqsillardan tashkil topgan murakkab ferment kompleksi tomonidan amalga oshiriladi. replikasoma ... Sintez uchun material hujayralar sitoplazmasida mavjud bo'lgan erkin nukleotidlardir. Replikatsiyaning biologik ma'nosi, odatda bo'linish paytida sodir bo'ladigan irsiy ma'lumotni ota-molekuladan qiziga aniq o'tkazishdadir. somatik hujayralar.
DNK molekulasi bir-birini to'ldiruvchi ikkita zanjirdan iborat. Bu zanjirlar zaif saqlanadi vodorod aloqalari fermentlar tomonidan parchalanishi mumkin. DNK molekulasi o'z-o'zidan ikkilanish (replikatsiya) qobiliyatiga ega va molekulaning har bir eski yarmida molekulaning yangi yarmi sintezlanadi.
Bundan tashqari, i-RNK molekulasi DNK molekulasida sintezlanishi mumkin, keyinchalik u DNKdan olingan ma'lumotni oqsil sintezi joyiga o'tkazadi.
Axborotni uzatish va oqsil sintezi matritsa printsipiga asoslanadi, bosmaxonadagi bosmaxonaning ishlashi bilan taqqoslanadi. DNKdan olingan ma'lumotlar ko'p marta ko'chiriladi. Agar nusxa ko'chirishda xatolar yuzaga kelsa, ular keyingi barcha nusxalarda takrorlanadi.

To'g'ri, DNK molekulasi tomonidan ma'lumotlarni nusxalashda ba'zi xatolar tuzatilishi mumkin - xatolarni bartaraf etish jarayoni deyiladi. kompensatsiyalar... Axborotni uzatish jarayonidagi reaktsiyalarning birinchisi DNK molekulasining replikatsiyasi va yangi DNK zanjirlarining sintezidir.
2. Transkripsiya (lot. transcriptio — qayta yozish) — barcha tirik hujayralarda uchraydigan DNK dan matritsa sifatida foydalanib, RNK sintezi jarayoni. Boshqacha qilib aytganda, bu genetik ma'lumotni DNKdan RNKga o'tkazishdir.
Transkripsiya DNKga bog'liq bo'lgan RNK polimeraza fermenti tomonidan katalizlanadi. RNK polimeraza DNK molekulasi bo'ylab 3 "→ 5" yo'nalishda harakat qiladi. Transkripsiya bosqichlardan iborat boshlanishi, cho'zilishi va tugashi ... Transkripsiya birligi operon bo'lib, DNK molekulasining fragmentidan iborat. promouter, transkripsiyalangan qism va terminator ... i-RNK bir zanjirdan iborat va i-RNK molekulasi sintezining boshlanishi va oxirini faollashtiruvchi ferment ishtirokida komplementarlik qoidasiga muvofiq DNKda sintezlanadi.
Tayyor i-RNK molekulasi sitoplazmaga ribosomalarga kiradi, bu erda polipeptid zanjirlarining sintezi sodir bo'ladi.
3. Translyatsiya (latdan. tarjima- transfer, harakat) - ribosoma tomonidan amalga oshiriladigan axborot (matritsa) RNK (mRNK, mRNK) matritsasidagi aminokislotalardan oqsil sintezi jarayoni. Boshqacha qilib aytganda, bu m-RNKning nukleotidlar ketma-ketligidagi ma'lumotlarni polipeptiddagi aminokislotalar ketma-ketligiga o'tkazish jarayonidir.

4. Teskari transkripsiya bir zanjirli RNKdan olingan ma'lumotlarga asoslangan ikki zanjirli DNK hosil bo'lish jarayonidir. Bu jarayon teskari transkripsiya deb ataladi, chunki bu holda genetik ma'lumotlarning uzatilishi transkripsiyaga nisbatan "teskari" yo'nalishda sodir bo'ladi. Teskari transkripsiya g'oyasi dastlab juda mashhur emas edi, chunki u markaziy dogmaga zid edi. molekulyar biologiya, bu DNKning RNKga transkripsiyasini va keyin oqsillarga aylanishini taklif qildi.
 Biroq, 1970 yilda Temin va Baltimor mustaqil ravishda ferment deb nomlangan fermentni topdilar teskari transkriptaza (teskari transkriptaza)
, va teskari transkripsiya imkoniyati nihoyat tasdiqlandi. 1975 yilda Temin va Baltimor mukofotlandi Nobel mukofoti fiziologiya va tibbiyot sohasida. Ba'zi viruslar (masalan, OIV infektsiyasini keltirib chiqaradigan odamning immunitet tanqisligi virusi) RNKni DNKga transkripsiya qilish qobiliyatiga ega. OIV DNKga singib ketgan RNK genomiga ega. Natijada, virusning DNKsi mezbon hujayraning genomi bilan birlashtirilishi mumkin. RNK dan DNK sintezi uchun mas'ul bo'lgan asosiy ferment deyiladi qaytish... Revertaza funktsiyalaridan biri yaratishdir komplementar DNK
(cDNK) virus genomidan. Bog'langan ribonukleaza fermenti RNKni parchalaydi, teskari transkriptaza esa DNK qo'sh spiralidan cDNKni sintez qiladi. cDNK integraza yordamida xost hujayra genomiga birlashtiriladi. Natija mezbon hujayra tomonidan virus oqsillarini sintezi yangi viruslarni hosil qiladi. OIV bo'lsa, T-limfotsitlarning apoptozi (hujayra o'limi) ham dasturlashtirilgan. Boshqa hollarda hujayra viruslarning tarqatuvchisi bo'lib qolishi mumkin.
Biroq, 1970 yilda Temin va Baltimor mustaqil ravishda ferment deb nomlangan fermentni topdilar teskari transkriptaza (teskari transkriptaza)
, va teskari transkripsiya imkoniyati nihoyat tasdiqlandi. 1975 yilda Temin va Baltimor mukofotlandi Nobel mukofoti fiziologiya va tibbiyot sohasida. Ba'zi viruslar (masalan, OIV infektsiyasini keltirib chiqaradigan odamning immunitet tanqisligi virusi) RNKni DNKga transkripsiya qilish qobiliyatiga ega. OIV DNKga singib ketgan RNK genomiga ega. Natijada, virusning DNKsi mezbon hujayraning genomi bilan birlashtirilishi mumkin. RNK dan DNK sintezi uchun mas'ul bo'lgan asosiy ferment deyiladi qaytish... Revertaza funktsiyalaridan biri yaratishdir komplementar DNK
(cDNK) virus genomidan. Bog'langan ribonukleaza fermenti RNKni parchalaydi, teskari transkriptaza esa DNK qo'sh spiralidan cDNKni sintez qiladi. cDNK integraza yordamida xost hujayra genomiga birlashtiriladi. Natija mezbon hujayra tomonidan virus oqsillarini sintezi yangi viruslarni hosil qiladi. OIV bo'lsa, T-limfotsitlarning apoptozi (hujayra o'limi) ham dasturlashtirilgan. Boshqa hollarda hujayra viruslarning tarqatuvchisi bo'lib qolishi mumkin.
Protein biosintezidagi matritsa reaktsiyalarining ketma-ketligini diagramma sifatida ko'rsatish mumkin.

Shunday qilib, oqsil biosintezi- Bu plastik metabolizm turlaridan biri bo'lib, uning davomida DNK genlarida kodlangan irsiy ma'lumot oqsil molekulalarida aminokislotalarning ma'lum bir ketma-ketligida amalga oshiriladi.
Protein molekulalari asosan polipeptid zanjirlari individual aminokislotalardan tashkil topgan. Ammo aminokislotalar o'z-o'zidan bir-biriga bog'lanish uchun etarlicha faol emas. Shuning uchun, bir-biri bilan bog'lanib, oqsil molekulasini hosil qilishdan oldin, aminokislotalar kerak faollashtirish ... Ushbu faollashuv maxsus fermentlar ta'sirida sodir bo'ladi.
Faollashuv natijasida aminokislota labil bo'ladi va xuddi shu ferment ta'sirida t- bilan bog'lanadi. RNK... Har bir aminokislota qat'iy o'ziga xos t-ga mos keladi. RNK, "o'z" aminokislotasini topadigan va olib boradi uni ribosomaga kiritadi.
Shuning uchun, har xil ular bilan birlashtirilgan faollashtirilgan aminokislotalar T- RNK... Ribosoma xuddi shunday konveyer unga kiradigan turli xil aminokislotalardan oqsil zanjirini yig'ish.
O'zining aminokislotasi "o'tirgan" t-RNK bilan bir vaqtda ribosoma " signal"Yadro tarkibidagi DNKdan. Ushbu signalga muvofiq, ribosomada ma'lum bir oqsil sintezlanadi.
DNKning oqsil sinteziga yo'naltiruvchi ta'siri to'g'ridan-to'g'ri emas, balki maxsus vositachi yordamida amalga oshiriladi - matritsa yoki xabarchi RNK (m-RNK yoki i-RNK), qaysi yadroga sintezlanadi e DNK ta'sirida, shuning uchun uning tarkibi DNK tarkibini aks ettiradi. RNK molekulasi DNK shaklidagi qolipga o'xshaydi. Sintezlangan i-RNK ribosomaga kiradi va xuddi shu tuzilishga o'tadi. reja- ribosomaga kiradigan faollashgan aminokislotalar ma'lum bir oqsilni sintez qilish uchun qanday tartibda bir-biri bilan birlashishi kerak. Aks holda, genetik ma'lumot DNKda kodlangan m-RNKga, keyin esa oqsilga o'tadi.
i-RNK molekulasi ribosomaga kiradi va tikuvlar uni. O'z ichiga olgan segment bu daqiqa ribosomada aniqlangan kodon (uchlik), mos tuzilma bilan juda aniq o'zaro ta'sir qiladi uchlik (antikodon) aminokislotalarni ribosomaga olib keladigan transport RNKsida.
Aminokislota bilan transport RNK ma'lum bir mRNK kodoniga mos keladi va bog‘laydi u bilan; keyingi, qo'shni saytga va-RNKga boshqa t-RNKni boshqa aminokislota bilan birlashtiradi va shunga o'xshash i-RNKning butun zanjiri o'qilguncha, barcha aminokislotalar tegishli tartibda bog'lanib, oqsil molekulasini hosil qilguncha davom etadi. Va polipeptid zanjirining ma'lum bir joyiga aminokislota etkazib beradigan t-RNK, aminokislotadan ozod qilinadi va ribosomani tark etadi.
Keyin yana sitoplazmada kerakli aminokislota unga biriktirilishi mumkin va u yana ribosomaga o'tadi. Oqsil sintezi jarayonida bir vaqtda bir emas, bir nechta ribosomalar - poliribosomalar ishtirok etadi.
Genetik ma'lumotni uzatishning asosiy bosqichlari:
1. i-RNK shablonidagi kabi DNKda sintez (transkripsiya)
2. m-RNK tarkibidagi dastur bo'yicha polipeptid zanjiri ribosomalarida sintez (translyatsiya)
.
Bosqichlar barcha tirik mavjudotlar uchun universaldir, lekin bu jarayonlarning vaqtinchalik va fazoviy munosabatlari pro- va eukariotlarda farqlanadi.
bor prokaryot DNK sitoplazmada bo'lgani uchun transkripsiya va tarjima bir vaqtning o'zida amalga oshirilishi mumkin. bor eukariotlar transkripsiya va translatsiya makon va vaqt bo'yicha qat'iy ravishda ajratiladi: yadroda turli RNKlarning sintezi sodir bo'ladi, shundan so'ng RNK molekulalari yadro membranasidan o'tib, yadrodan chiqib ketishi kerak. Keyinchalik, sitoplazmada RNKlar oqsil sintezi joyiga ko'chiriladi.
5-ma'ruza. Genetik kod
Kontseptsiyaning ta'rifi
Genetik kod - bu DNKdagi nukleotidlarning joylashish ketma-ketligidan foydalangan holda oqsillardagi aminokislotalarning ketma-ketligi haqidagi ma'lumotlarni yozib olish tizimi.
DNK oqsil sintezida bevosita ishtirok etmaganligi sababli kod RNK tilida yozilgan. RNK tarkibida timin o'rniga urasil mavjud.
Genetik kodning xususiyatlari
1. Uchlik
Har bir aminokislota 3 ta nukleotidlar ketma-ketligi sifatida kodlangan.
Ta'rif: triplet yoki kodon - bitta aminokislotalarni kodlaydigan uchta nukleotidlar ketma-ketligi.
Kod singlet bo'lishi mumkin emas, chunki 4 (DNKdagi turli nukleotidlar soni) 20 dan kam. Kod dublet bo'lishi mumkin emas, chunki 16 (4 ta nukleotidning 2 ga birikmalari va almashinishlari soni) 20 dan kam. Kod uchlik bo'lishi mumkin, chunki 64 (4 dan 3 gacha kombinatsiyalar va almashtirishlar soni) 20 dan ortiq.
2. Degeneratsiya.
Barcha aminokislotalar, metionin va triptofandan tashqari, bir nechta tripletlar bilan kodlangan:
2 AK 1 triplet = 2.
9 AK 2 uchlik = 18.
1 AK 3 uchlik = 3.
5 AK 4 uchlik = 20.
3 AK 6 uchlik = 18.
Jami 61 triplet 20 ta aminokislotalarni kodlaydi.
3. Genlararo tinish belgilarining mavjudligi.
Ta'rifi:
Gen bir polipeptid zanjiri yoki bitta molekulani kodlaydigan DNK qismidir tPHK, rRNK yokisPHK.
GenlartPHK, rPHK, sPHKoqsillar kodlanmaydi.
Polipeptidni kodlaydigan har bir genning oxirida RNK to'xtash kodonlarini yoki to'xtash signallarini kodlaydigan 3 ta tripletdan kamida bittasi joylashgan. mRNKda ular quyidagicha ko'rinadi: UAA, UAG, UGA ... Ular eshittirishni tugatadilar (tugaydilar).
An'anaviy ravishda kodon tinish belgilariga ham tegishli AVG - yetakchi ketma-ketligidan keyingi birinchi. (8-ma'ruzaga qarang) U bosh harf vazifasini bajaradi. Bu holatda u formilmetioninni (prokaryotlarda) kodlaydi.
4. Aniqlik.
Har bir triplet faqat bitta aminokislotani kodlaydi yoki tarjima terminatoridir.
Istisno - bu kodon AVG ... Prokaryotlarda, birinchi o'rinda (katta harf) u formilmetioninni va boshqa har qanday holatda - metioninni kodlaydi.
5. Kompaktlik yoki intragenik tinish belgilarining yo'qligi.
Gen ichida har bir nukleotid ma'no kodonining bir qismidir.
1961 yilda Seymur Benzer va Frensis Krik kod uchlik va ixcham ekanligini eksperimental tarzda isbotladilar.
Tajribaning mohiyati: "+" mutatsiyasi - bitta nukleotidning kiritilishi. "-" mutatsiyasi - bu bitta nukleotidning yo'qolishi. Genning boshida bitta "+" yoki "-" mutatsiya butun genni buzadi. Ikki tomonlama "+" yoki "-" mutatsiya ham butun genni buzadi.
Genning boshida uch karra "+" yoki "-" mutatsiya uning faqat bir qismini buzadi. To'rtlik + yoki - mutatsiya yana butun genni buzadi.
Tajriba buni isbotlaydi kod murakkab va gen ichida tinish belgilari yo'q. Tajriba ikkita qo'shni fag genida o'tkazildi va bundan tashqari, genlar orasidagi tinish belgilarining mavjudligi.
6. Ko'p qirralilik.
Er yuzida yashovchi barcha mavjudotlar uchun genetik kod bir xil.
1979 yilda Burrell ochildi ideal inson mitoxondriya kodi.
Ta'rifi:
"Ideal" genetik kod bo'lib, unda kvazi-dubl kodning degeneratsiyasi qoidasi bajariladi: Agar birinchi ikkita nukleotid ikkita tripletga to'g'ri kelsa va uchinchi nukleotidlar bir xil sinfga tegishli bo'lsa (ikkalasi ham purinlar yoki ikkalasi ham pirimidinlar). , keyin bu uchlik bir xil aminokislotalarni kodlaydi ...
Umumiy kodda ushbu qoidadan ikkita istisno mavjud. Umumjahondagi ideal koddan ikkala og'ish ham asosiy nuqtalarga tegishli: oqsil sintezining boshlanishi va oxiri:
Kodon | Universal kod | Mitoxondriyal kodlar |
|||
Umurtqali hayvonlar | Umurtqasizlar | Xamirturush | O'simliklar |
||
STOP | STOP |
||||
UA bilan | |||||
A G A | STOP | ||||
STOP | 230 ta almashtirish kodlangan aminokislota sinfini o'zgartirmaydi. yirtib tashlash qobiliyatiga. 1956 yilda Georgiy Gamov bir-biriga o'xshash kodning variantini taklif qildi. Gamow kodiga ko'ra, gendagi uchinchidan boshlab har bir nukleotid 3 ta kodonga kiradi. Genetik kod dekodlanganda, u bir-biriga mos kelmasligi ma'lum bo'ldi, ya'ni. har bir nukleotid faqat bitta kodonga kiradi. Bir-biriga o'xshash genetik kodning afzalliklari: ixchamlik, oqsil tuzilishining nukleotidlarni kiritish yoki yo'q qilishga kamroq bog'liqligi. Kamchilik: oqsil tuzilishining nukleotidlarni almashtirishga yuqori bog'liqligi va qo'shnilarga cheklov. 1976 yilda phX174 fagining DNKsi ketma-ketlashtirildi. U 5375 nukleotiddan iborat bir zanjirli dumaloq DNKga ega. Ma'lumki, fag 9 ta oqsilni kodlaydi. Ulardan 6 tasi uchun birin-ketin joylashgan genlar aniqlangan. Ma'lum bo'lishicha, bir-biriga o'xshashlik bor. Gen E butunlay gen ichida D ... Uning boshlang'ich kodoni bir nukleotid o'qilishining siljishi natijasida paydo bo'ladi. Gen J gen tugagan joydan boshlanadi D ... Genni boshlash kodoni J genning tugatish kodoni bilan ustma-ust tushadi D ikki nukleotidning siljishi natijasida. Konstruksiya uchga karrali bo'lmagan nukleotidlar soni tomonidan "o'qish ramkasining siljishi" deb ataladi. Bugungi kunga kelib, bir-biriga o'xshashlik faqat bir nechta faglar uchun ko'rsatilgan. DNK ma'lumot qobiliyati Yer yuzida 6 milliard odam yashaydi. Ular haqida irsiy ma'lumotlar 4x10 13 kitob sahifalari. Bu sahifalar NDUning 6 ta binosi hajmini egallaydi. 6x10 9 spermatozoidlar oyukning yarmini egallaydi. Ularning DNKsi to'rtdan bir qismidan kamroq qismini egallaydi. | ||||
- nuklein kislota molekulalaridagi irsiy ma'lumotlarni nukleotidlar ketma-ketligi ko'rinishida qayd etishning yagona tizimi. Genetik kod azotli asoslarda farq qiluvchi faqat to'rtta harf - nukleotidlardan iborat alifbodan foydalanishga asoslangan: A, T, G, C.
Genetik kodning asosiy xususiyatlari quyidagilardan iborat:
1. Genetik kod uchlikdir. Triplet (kodon) - bitta aminokislotalarni kodlaydigan uchta nukleotidlar ketma-ketligi. Proteinlar 20 ta aminokislotadan iborat bo'lganligi sababli, ularning har birini bitta nukleotid bilan kodlash mumkin emasligi aniq (chunki DNKda faqat to'rt turdagi nukleotidlar mavjud, bu holda 16 ta aminokislotalar kodlanmagan holda qoladi). Aminokislotalarni kodlash uchun ikkita nukleotid ham yo'q, chunki bu holda faqat 16 ta aminokislotani kodlash mumkin. Bu shuni anglatadiki, bitta aminokislotani kodlaydigan nukleotidlarning eng kichik soni uchta bo'lib chiqadi. (Bu holda nukleotidlarning mumkin bo'lgan tripletlari soni 4 3 = 64).
2. Kodning ortiqcha bo'lishi (degeneratsiyasi) uning uchligining oqibati bo'lib, bitta aminokislota bir nechta tripletlar bilan kodlanishi mumkinligini anglatadi (chunki 20 ta aminokislota va 64 ta triplet mavjud). Istisnolar faqat bitta triplet tomonidan kodlangan metionin va triptofandir. Bundan tashqari, ba'zi tripletlar o'ziga xos funktsiyalarga ega. Shunday qilib, mRNK molekulasida ulardan uchtasi, UAA, UAH va UGA, tugatish kodonlari, ya'ni polipeptid zanjiri sintezini to'xtatuvchi signallardir. DNK zanjirining boshida joylashgan metionin (AUG) ga mos keladigan triplet aminokislotalarni kodlamaydi, balki o'qishni boshlash (qo'zg'atish) funktsiyasini bajaradi.
3. Ortiqchalik bilan bir vaqtda kod noaniqlik xususiyatiga ega, ya'ni har bir kodon faqat bitta o'ziga xos aminokislotaga mos keladi.
4. Kod kollinear; gendagi nukleotidlar ketma-ketligi oqsildagi aminokislotalar ketma-ketligiga to'liq mos keladi.
5. Genetik kod bir-birining ustiga chiqmaydi va ixchamdir, ya'ni unda "tinish belgilari" mavjud emas. Bu shuni anglatadiki, o'qish jarayoni ustunlar (uchlik) bir-birining ustiga tushishiga yo'l qo'ymaydi va ma'lum bir kodondan boshlab, o'qish doimiy ravishda, uchlikdan keyin uch marta, to'xtash signallarigacha (tugatish kodonlari) davom etadi. Masalan, mRNKda AUGGUGTSUUAAUGUG azotli asoslarning quyidagi ketma-ketligi faqat shunday tripletlar tomonidan o'qiladi: AUG, UGG, GGU, GUG va boshqalar emas, balki AUG, GUG, CUU, AAU, GUG yoki AUG, GGU, UGC, CUU va boshqalar yoki boshqa usulda (masalan, AUG kodon, tinish belgisi G, UGC kodon, U tinish belgisi va boshqalar).
6. Genetik kod universaldir, ya'ni barcha organizmlarning yadro genlari oqsillar haqidagi ma'lumotlarni, bu organizmlarning tashkiliy darajasi va sistematik holatidan qat'i nazar, bir xil tarzda kodlaydi.
Genetik kod- nuklein kislota molekulalaridagi irsiy ma'lumotlarni nukleotidlar ketma-ketligi ko'rinishida qayd etishning yagona tizimi. Genetik kod DNK nukleotidlariga mos keladigan faqat to'rtta A, T, C, G harflaridan iborat alifbodan foydalanishga asoslangan. Hammasi bo'lib 20 turdagi aminokislotalar mavjud. 64 ta kodondan uchtasi - UAA, UAG, UGA - aminokislotalarni kodlamaydi, ular bema'ni kodonlar deb ataladi va ular tinish belgilari sifatida ishlaydi. Kodon (trinukleotidni kodlovchi) genetik kodning birligi, DNK yoki RNKdagi nukleotid qoldiqlarining uchligi (uchlik) bo'lib, bitta aminokislota kiritilishini kodlaydi. Genlarning o'zi oqsil sintezida ishtirok etmaydi. Gen va oqsil o'rtasidagi vositachi mRNKdir. Genetik kodning tuzilishi uning triplet bo'lishi, ya'ni kodon deb ataladigan DNKning azotli asoslarining tripletlari (uchliklari) dan iboratligi bilan tavsiflanadi. 64 dan
Gen xususiyatlari. kod
1) Triplet: bitta aminokislota uchta nukleotid bilan kodlangan. DNKdagi bu 3 ta nukleotid
triplet, mRNKda - kodon, tRNKda - antikodon deb ataladi.
2) Ortiqchalik (degeneratsiya): atigi 20 ta aminokislota va aminokislotalarni kodlaydigan tripletlar 61, shuning uchun har bir aminokislota bir nechta tripletlar bilan kodlanadi.
3) Aniqlik: har bir triplet (kodon) faqat bitta aminokislotani kodlaydi.
4) Ko'p qirralilik: genetik kod Yerdagi barcha tirik organizmlar uchun bir xil.
5.) o'qishda kodonlarning uzluksizligi va izchilligi. Bu shuni anglatadiki, nukleotidlar ketma-ketligi bo'shliqlarsiz uch marta o'qiladi, qo'shni tripletlar esa bir-birining ustiga chiqmaydi.
88. Irsiyat va o'zgaruvchanlik tirik mavjudotlarning asosiy xususiyatlari. Darvinning irsiyat va o'zgaruvchanlik hodisalarini tushunishi.
Irsiyat ular barcha organizmlarning umumiy mulkini ota-onadan naslga xos xususiyatlarni saqlash va etkazish deb atashadi. Irsiyat- bu organizmlarning bu jarayonda rivojlangan o'xshash turdagi metabolizmni avlodlarda ko'paytirish xususiyatidir. tarixiy rivojlanish turlari va ma'lum sharoitlarda o'zini namoyon qiladi tashqi muhit.
O'zgaruvchanlik Bir xil turdagi individlar o'rtasida sifat farqlarining paydo bo'lish jarayoni mavjud bo'lib, u faqat bitta fenotipning tashqi muhit ta'siri ostida o'zgarishi yoki kombinatsiyalar, rekombinatsiyalar va mutatsiyalar natijasida genetik jihatdan aniqlangan irsiy o'zgarishlarda ifodalanadi. bir qator ketma-ket avlodlar va populyatsiyalarda uchraydi.
Darvinning irsiyat va o'zgaruvchanlik haqidagi tushunchasi.
Irsiyat ostida Darvin organizmlarning avlodlarida o'z turlarini, nav va individual xususiyatlarini saqlab qolish qobiliyatini tushundi. Bu xususiyat yaxshi ma'lum bo'lgan va irsiy o'zgaruvchanlikni ifodalagan. Darvin evolyutsiya jarayonida irsiyatning ahamiyatini batafsil tahlil qildi. U birinchi avlod duragaylarining bir xilligi va ikkinchi avlodda belgilarning bo'linishi holatlariga e'tibor qaratdi, u jins bilan bog'liq irsiyat, gibrid atavizmlar va irsiyatning bir qator boshqa hodisalaridan xabardor edi.
O'zgaruvchanlik. Hayvonlarning ko'p zotlari va o'simliklarning navlarini taqqoslab, Darvin hayvonlar va o'simliklarning har qanday turida va madaniyatda har qanday nav va zotda bir xil shaxslar mavjud emasligini ta'kidladi. Darvin oʻzgaruvchanlik barcha hayvonlar va oʻsimliklarga xos degan xulosaga keldi.
Hayvonlarning o'zgaruvchanligi haqidagi materialni tahlil qilar ekan, olim o'zgaruvchanlikni keltirib chiqarish uchun qamoqda saqlash sharoitlarining har qanday o'zgarishi etarli ekanligini ta'kidladi. Shunday qilib, Darvin o'zgaruvchanlikni organizmlarning atrof-muhit sharoitlari ta'sirida yangi belgilarga ega bo'lish qobiliyati deb tushundi. U o'zgaruvchanlikning quyidagi shakllarini ajratdi:
Maxsus (guruh) o'zgaruvchanlik(hozir chaqiriladi modifikatsiya) - ma'lum sharoitlarning ta'siri tufayli naslning barcha individlarida bir yo'nalishda o'xshash o'zgarish. Ba'zi o'zgarishlar odatda irsiy bo'lmaydi.
Noaniq individual o'zgaruvchanlik(hozir chaqiriladi genotipik) - bir xil sharoitda mavjud bo'lgan bir individ boshqalardan ajralib turadigan bir xil tur, nav, zotning individlarida har xil ahamiyatsiz farqlarning paydo bo'lishi. Bunday ko'p yo'nalishli o'zgaruvchanlik mavjudlik shartlarining har bir alohida shaxsga noaniq ta'sirining natijasidir.
Korrelyativ(yoki nisbiy) o'zgaruvchanlik. Darvin organizmni alohida qismlari bir-biri bilan chambarchas bog'langan yaxlit tizim sifatida tushundi. Shuning uchun bir qismning tuzilishi yoki funktsiyasining o'zgarishi ko'pincha boshqa yoki boshqalarning o'zgarishiga olib keladi. Bunday o'zgaruvchanlikka misol qilib, ishlaydigan mushakning rivojlanishi va u yopishgan suyakda tepalikning shakllanishi o'rtasidagi bog'liqlikdir. Ko'p suvli qushlarda bo'yin uzunligi va oyoq-qo'l uzunligi o'rtasida bog'liqlik mavjud: uzun bo'yinli qushlarning ham uzun oyoq-qo'llari bor.
Kompensatsion o'zgaruvchanlik shundan iboratki, ba'zi organlar yoki funktsiyalarning rivojlanishi ko'pincha boshqalarning zulmiga sabab bo'ladi, ya'ni teskari korrelyatsiya, masalan, qoramolning sut mahsuldorligi va go'shtliligi o'rtasida.
89. Modifikatsiyaning o‘zgaruvchanligi. Genetik jihatdan aniqlangan belgilarning reaktsiya tezligi. Fenokopiyalar.
Fenotipik o'zgaruvchanlik rivojlanish sharoitlari yoki atrof-muhit omillari ta'sirida yuzaga keladigan bevosita belgilar holatidagi o'zgarishlarni qamrab oladi. Modifikatsiyaning o'zgaruvchanligi diapazoni oddiy javob bilan cheklangan. Xususiyatdagi xos modifikatsiyadagi o’zgarish irsiy xususiyatga ega bo’lmaydi, lekin modifikatsiya o’zgaruvchanligi diapazoni irsiyat bilan belgilanadi.Bunda irsiy material o’zgarishda ishtirok etmaydi.
Reaktsiya tezligi- bu belgining modifikatsiya o'zgaruvchanligi chegarasi. Reaktsiya tezligi meros bo'lib o'tadi, lekin modifikatsiyalarning o'zi emas, ya'ni. xususiyatni rivojlantirish qobiliyati va uning namoyon bo'lish shakli atrof-muhit sharoitlariga bog'liq. Reaksiya tezligi genotipning o'ziga xos miqdoriy va sifat ko'rsatkichidir. Keng reaksiya tezligi, tor () va bir ma'noli tezligi bo'lgan belgilar mavjud. Reaktsiya tezligi har bir tur (pastki va yuqori) uchun chegaralari yoki chegaralari bor - masalan, oziqlantirishning ko'payishi hayvonning vaznining oshishiga olib keladi, ammo u ma'lum bir tur yoki zotga xos bo'lgan reaktsiya tezligi doirasida bo'ladi. Reaktsiya tezligi genetik jihatdan aniqlanadi va meros bo'lib o'tadi. Turli belgilar uchun reaktsiya normasining chegaralari juda farq qiladi. Masalan, sut mahsuldorligi, donli ekinlarning mahsuldorligi va boshqa ko'plab miqdoriy belgilar reaktsiya tezligi uchun keng chegaralarga ega, tor chegaralar ko'pchilik hayvonlarning rang intensivligi va boshqa ko'plab sifat belgilaridir. Ba'zilar ta'sirida zararli omillar, evolyutsiya jarayonida odam duch kelmaydigan, reaktsiya me'yorlarini belgilaydigan o'zgaruvchanlikni o'zgartirish imkoniyati istisno qilinadi.
Fenokopiyalar- atrof-muhitning noqulay omillari ta'sirida fenotipdagi o'zgarishlar, mutatsiyalarga o'xshash namoyon bo'lishi. Olingan fenotipik o'zgarishlar meros qilib olinmaydi. Fenokopiyaning paydo bo'lishi ma'lum bir cheklangan rivojlanish bosqichiga tashqi sharoitlarning ta'siri bilan bog'liqligi aniqlandi. Bundan tashqari, xuddi shu agent, qaysi fazada harakat qilishiga qarab, turli mutatsiyalarni nusxalashi mumkin yoki bir bosqich bir agentga, ikkinchisi boshqasiga reaksiyaga kirishadi. Xuddi shu fenokopiyani qo'zg'atish uchun turli xil vositalardan foydalanish mumkin, bu o'zgarish natijasi va ta'sir etuvchi omil o'rtasida hech qanday bog'liqlik yo'qligini ko'rsatadi. Rivojlanishning eng murakkab genetik kasalliklarini ko'paytirish nisbatan oson, belgilarni esa nusxalash ancha qiyin.
90. Modifikatsiyaning moslashish xususiyati. Shaxs kamoloti, ta'lim va tarbiyasida irsiyat va muhitning o'rni.
Modifikatsiyaning o'zgaruvchanligi yashash sharoitlariga mos keladi va moslashuvchan xususiyatga ega. O'simliklar va hayvonlarning o'sishi, ularning massasi, rangi va boshqalar kabi xususiyatlar modifikatsiyaning o'zgaruvchanligiga bog'liq. Modifikatsion o'zgarishlarning ko'rinishi atrof-muhit sharoitlari rivojlanayotgan organizmda sodir bo'ladigan fermentativ reaktsiyalarga ta'sir qilishi va ma'lum darajada uning yo'nalishini o'zgartirishi bilan bog'liq.
Irsiy ma'lumotlarning fenotipik namoyon bo'lishi atrof-muhit sharoitlari bilan o'zgartirilishi mumkinligi sababli, faqat reaktsiya normasi deb ataladigan ma'lum chegaralarda ularning hosil bo'lish imkoniyati organizm genotipida dasturlashtirilgan. Reaktsiya tezligi ma'lum bir genotip uchun ruxsat etilgan belgining o'zgaruvchan o'zgaruvchanligi chegaralarini ifodalaydi.
Turli sharoitlarda genotipni amalga oshirish jarayonida belgining jiddiyligi ekspressivlik deb ataladi. Bu xususiyatning normal reaktsiya oralig'ida o'zgaruvchanligi bilan bog'liq.
Xuddi shu xususiyat ba'zi organizmlarda paydo bo'lishi mumkin va bir xil genga ega bo'lgan boshqalarida yo'q. Genning fenotipik namoyon bo'lishining miqdoriy ko'rsatkichi penetratsiya deb ataladi.
Ekspressivlik va penetratsiya tabiiy tanlanish tomonidan qo'llab-quvvatlanadi. Odamlarda irsiyatni o'rganishda ikkala naqshni ham yodda tutish kerak. Atrof-muhit sharoitlarini o'zgartirish orqali penetratsiya va ekspressivlikka ta'sir qilish mumkin. Bitta va bir xil genotip turli fenotiplarning rivojlanishining manbai bo'lishi mumkinligi tibbiyot uchun juda muhimdir. Bu shuni anglatadiki, yuklangan kishi o'zini namoyon qilishi shart emas. Ko'p narsa odamning sharoitlariga bog'liq. Ba'zi hollarda kasallik irsiy ma'lumotlarning fenotipik namoyon bo'lishi sifatida dietaga rioya qilish yoki dori-darmonlarni qabul qilish orqali oldini olish mumkin. Irsiy ma'lumotlarning amalga oshirilishi atrof-muhitga bog'liq.Tarixiy shakllangan genotip asosida shakllangan modifikatsiyalar odatda adaptivdir, chunki ular doimo javob reaktsiyalari natijasidir. rivojlanayotgan organizm ustida ekologik omillar... Mutatsion o'zgarishlarning tabiati har xil: ular DNK molekulasi strukturasidagi o'zgarishlar natijasidir, bu esa ilgari tashkil etilgan oqsil sintezi jarayonining buzilishiga olib keladi. sichqonlar yuqori harorat sharoitida saqlansa, ular dumlari cho'zilgan va quloqlari kattalashgan nasl tug'adilar. Ushbu modifikatsiya tabiatda adaptivdir, chunki chiqadigan qismlar (dum va quloqlar) tanada termoregulyatsiya rolini o'ynaydi: ularning sirtining oshishi issiqlik o'tkazuvchanligini oshirishga imkon beradi.
Insonning genetik salohiyati vaqt bilan cheklangan va juda qattiq. Agar siz erta sotsializatsiya muddatini o'tkazib yuborsangiz, u yo'qoladi, amalga oshirishga vaqt topolmaydi. Ushbu bayonotning yorqin misoli - chaqaloqlar o'rmonga tushib, bir necha yil hayvonlar orasida o'tkazgan ko'plab holatlardir. Insoniyat jamiyatiga qaytganlaridan so'ng, ular yo'qotilgan vaqtni to'liq qoplay olmadilar: nutqni o'zlashtira olmadilar, inson faoliyatining ancha murakkab ko'nikmalariga ega bo'ldilar, insonning aqliy funktsiyalari yomon rivojlangan. Bu insonning xulq-atvori va faoliyatining xarakterli xususiyatlari faqat ijtimoiy meros orqali, faqat ijtimoiy dasturni ta'lim va tarbiya jarayonida o'tkazish orqali ega bo'lishidan dalolat beradi.
Turli xil muhitda bo'lgan bir xil genotiplar (bir xil egizaklarda) turli xil fenotiplarni berishi mumkin. Barcha ta'sir omillarini hisobga olgan holda, inson fenotipini bir nechta elementlardan iborat sifatida ko'rsatish mumkin.
Bularga quyidagilar kiradi: genlarda kodlangan biologik moyilliklar; atrof-muhit (ijtimoiy va tabiiy); shaxsning faoliyati; aql (ong, fikrlash).
Inson rivojlanishida irsiyat va atrof-muhitning o'zaro ta'siri uning hayoti davomida muhim rol o'ynaydi. Ammo u organizmning shakllanish davrlarida alohida ahamiyatga ega bo'ladi: embrion, ko'krak, bola, o'smir va yosh. Aynan shu davrda organizmning rivojlanishi va shaxs shakllanishining intensiv jarayoni kuzatildi.
Irsiyat organizmning nimaga aylanishi mumkinligini aniqlaydi, lekin inson bir vaqtning o'zida ikkala omil - irsiyat va atrof-muhit ta'siri ostida rivojlanadi. Bugungi kunda insonning moslashuvi irsiyatning ikkita dasturi ta'siri ostida amalga oshirilishi umumiy qabul qilinmoqda: biologik va ijtimoiy. Har qanday shaxsning barcha belgilari va xususiyatlari uning genotipi va atrof-muhitining o'zaro ta'siri natijasidir. Binobarin, har bir shaxs ham tabiatning bir qismi, ham ijtimoiy taraqqiyot mahsulidir.
91. Kombinativ o‘zgaruvchanlik. Odamlarning genotipik xilma-xilligini ta'minlashda kombinativ o'zgaruvchanlikning ahamiyati: Nikoh tizimlari. Oilaning tibbiy va genetik jihatlari.
Kombinativ o'zgaruvchanlik genotipdagi genlarning yangi birikmalarini olish bilan bog'liq. Bunga uchta jarayon natijasida erishiladi: a) meioz davrida xromosomalarning mustaqil divergentsiyasi; b) urug'lantirish paytida ularning tasodifiy birikmasi; c) Krossover tufayli genlarning rekombinatsiyasi. Irsiy omillarning (genlarning) o'zi o'zgarmaydi, lekin ularning yangi birikmalari paydo bo'ladi, bu esa boshqa genotipik va fenotipik xususiyatlarga ega bo'lgan organizmlarning paydo bo'lishiga olib keladi. Kombinativ o'zgaruvchanlik tufayli ega bo'lgan naslda turli xil genotiplar yaratiladi katta ahamiyatga ega evolyutsiya jarayoni uchun quyidagilar tufayli: 1)
evolyutsiya jarayoni uchun materiallarning xilma-xilligi shaxslarning hayotiyligini kamaytirmasdan ortadi; 2)
organizmlarning o'zgaruvchan atrof-muhit sharoitlariga moslashish imkoniyatlari kengayadi va shu bilan organizmlar guruhining (populyar, turlar) umuman yashashini ta'minlaydi.
Odamlarda, populyatsiyalarda allellarning tarkibi va chastotasi ko'p jihatdan nikoh turlariga bog'liq. Shu munosabat bilan nikoh turlari va ularning tibbiy-genetik oqibatlarini o'rganish katta ahamiyatga ega.
Nikoh quyidagilar bo'lishi mumkin: saylov, beg'araz.
Farq qilmaslik panmix nikohlarini o'z ichiga oladi. Panmiksiya(yunoncha nixis - aralash) - turli genotipli odamlar o'rtasidagi yarim nikoh.
Saylov asosidagi nikohlar: 1. Outbreding- oldindan belgilangan genotip bo'yicha oilaviy aloqalarga ega bo'lmagan odamlar o'rtasidagi nikohlar; 2. Inbreeding- qarindoshlar o'rtasidagi nikohlar; 3.Ijobiy-assortativ- fenotiplari o'xshash bo'lgan shaxslar o'rtasidagi nikohlar (kar va soqov, past bo'ylilar bilan, baland bo'ylilar bilan, zaif aqllilar bilan zaif va boshqalar). 4.Salbiy-assortativ-fenotiplari bir-biriga o'xshamaydigan odamlar o'rtasidagi nikohlar (kar-soqov-normal; kalta-bo'yli; normal - sepkilli va boshqalar). 4 insest- yaqin qarindoshlar o'rtasidagi nikohlar (aka va opa-singillar o'rtasidagi).
Ko'pgina mamlakatlarda qarindoshlar va qarindoshlar o'rtasidagi nikoh noqonuniy hisoblanadi. Afsuski, qarindoshlar o'rtasidagi nikohlar yuqori bo'lgan hududlar mavjud. Yaqin vaqtgacha O‘rta Osiyoning ayrim hududlarida qarindosh-urug‘lar bilan nikoh qurishlar soni 13-15 foizga yetgan.
Tibbiy va genetik ahamiyati Inbred nikohlar juda salbiy. Bunday nikohlar bilan gomozigotizatsiya kuzatiladi, otosomal retsessiv kasalliklarning chastotasi 1,5-2 barobar ortadi. Inbred populyatsiyalar inbred depressiya bilan tavsiflanadi, ya'ni. chastotasi keskin ortadi, kiruvchi retsessiv allellarning chastotasi oshadi va chaqaloqlar o'limi ortadi. Ijobiy-assortativ nikohlar ham shunga o'xshash hodisalarga olib keladi. Outbreeding genetik jihatdan ijobiydir. Bunday nikohlar bilan heterozigotizatsiya kuzatiladi.
92. Mutatsion o'zgaruvchanlik, irsiy materialning shikastlanishining o'zgarishlar darajasiga ko'ra mutatsiyalarning tasnifi. Jinsiy va somatik hujayralardagi mutatsiyalar.
Mutatsiya ko'payish tuzilmalarining qayta tashkil etilishi, uning genetik apparatining o'zgarishi tufayli o'zgarish deyiladi. Mutatsiyalar spazmodik tarzda sodir bo'ladi va meros qilib olinadi. Irsiy materialning o'zgarish darajasiga qarab, barcha mutatsiyalar bo'linadi gen, xromosoma va genomik.
Gen mutatsiyalari, yoki transgenatsiyalar genning tuzilishiga ta'sir qiladi. Mutatsiyalar DNK molekulasining turli uzunlikdagi bo'limlarini o'zgartirishi mumkin. Mutatsiya paydo bo'lishiga olib keladigan o'zgarishlarning eng kichik joyiga muton deyiladi. Bu faqat bir nechta nukleotidlar bo'lishi mumkin. DNKdagi nukleotidlar ketma-ketligining o'zgarishi tripletlar ketma-ketligining o'zgarishini va oxir-oqibat, oqsil sintezi dasturini belgilaydi. Shuni esda tutish kerakki, DNK tuzilishidagi buzilishlar faqat ta'mirlash amalga oshirilmaganda mutatsiyaga olib keladi.
Xromosoma mutatsiyalari, xromosomalarning qayta tuzilishi yoki aberratsiyasi xromosomalarning irsiy materialining sonining o'zgarishi yoki qayta taqsimlanishidan iborat.
Qayta qurish quyidagilarga bo'linadi nutrikromosomali va xromosomalararo... Xromosoma ichidagi qayta tuzilishlar xromosomaning bir qismini yo'qotish (yo'q qilish), uning ba'zi bo'limlarini ikki baravar oshirish yoki ko'paytirish (ko'paytirish), xromosoma bo'lagining genlar ketma-ketligining o'zgarishi (inversiya) bilan 180 ° ga aylanishidan iborat.
Genomik mutatsiyalar xromosomalar sonining o'zgarishi bilan bog'liq. Genomik mutatsiyalarga anevloidiya, gaploidiya va poliploidiya kiradi.
Anevloidiya individual xromosomalar sonining o'zgarishi deyiladi - yo'qligi (monosomiya) yoki qo'shimcha (trisomiya, tetrasomiya, umumiy holda, polisomiya) xromosomalarning mavjudligi, ya'ni muvozanatsiz xromosomalar to'plami. Xromosomalar soni o'zgargan hujayralar mitoz yoki meyoz jarayonlaridagi buzilishlar natijasida paydo bo'ladi, bu bilan bog'liq holda mitotik va meiotik anevlodiya farqlanadi. Somatik hujayralar xromosomalari sonining diploid bilan solishtirganda bir necha marta kamayishi deyiladi. gaploid... Somatik hujayralar xromosoma to'plamlari sonining diploidga nisbatan bir necha marta ko'payishi deyiladi. poliploidiya.
Ro'yxatdagi turlar mutatsiyalar jinsiy hujayralarda ham, somatik hujayralarda ham uchraydi. Jinsiy hujayralarda yuzaga keladigan mutatsiyalar deyiladi generativ... Ular keyingi avlodlarga o'tadi.
Organizmning individual rivojlanishining u yoki bu bosqichida tana hujayralarida yuzaga keladigan mutatsiyalar deyiladi somatik... Bunday mutatsiyalar faqat u sodir bo'lgan hujayraning avlodlari tomonidan meros qilib olinadi.
93. Gen mutatsiyalari, paydo bo'lish molekulyar mexanizmlari, tabiatdagi mutatsiyalarning chastotasi. Mutatsiyaga qarshi biologik mexanizmlar.
Zamonaviy genetika buni ta'kidlaydi gen mutatsiyalari genlarning kimyoviy tuzilishini o'zgartirishdan iborat. Xususan, gen mutatsiyalari asosiy juftliklarni almashtirish, kiritish, tushirish va yo'qotishdir. DNK molekulasining o'zgarishi mutatsiyaga olib keladigan eng kichik qismiga muton deyiladi. U bir juft nukleotidga teng.
Gen mutatsiyalarining bir nechta tasnifi mavjud ... O'z-o'zidan(o'z-o'zidan) - atrof-muhitdagi har qanday fizik yoki kimyoviy omil bilan bevosita bog'liqlikdan tashqari yuzaga keladigan mutatsiya.
Agar mutatsiyalar ataylab, organizmga ma'lum tabiat omillari ta'sirida yuzaga kelgan bo'lsa, ular deyiladi. qo'zg'atilgan... Mutatsiyani qo'zg'atuvchi vosita deyiladi mutagen.
Mutagenlarning tabiati xilma-xildir jismoniy omillar, kimyoviy birikmalar... Ba'zi biologik ob'ektlar - viruslar, oddiylar, gelmintlar inson tanasiga tushganda mutagen ta'siri aniqlangan.
Dominant va retsessiv mutatsiyalar natijasida fenotipda dominant va retsessiv o'zgargan belgilar paydo bo'ladi. Dominant Mutatsiyalar fenotipda birinchi avlodda paydo bo'ladi. Resessiv mutatsiyalar harakatdan geterozigotalarda yashiringan tabiiy tanlanish, shuning uchun ular turlarning genofondida ko'p miqdorda to'planadi.
Mutatsiya jarayoni intensivligining ko'rsatkichi mutatsiya chastotasi bo'lib, u har bir genom uchun o'rtacha yoki alohida lokuslar uchun alohida hisoblanadi. O'rtacha mutatsiya chastotasi tirik mavjudotlarning keng doirasi (bakteriyalardan odamlargacha) bilan taqqoslanadi va morfofiziologik tashkilotning darajasi va turiga bog'liq emas. Bir avlodda 1 lokusga 10 -4 - 10 -6 mutatsiyaga teng.
Mutatsiyaga qarshi mexanizmlar.
Eukaryotik somatik hujayralarning diploid karyotipidagi xromosomalar juftligi gen mutatsiyalarining salbiy ta'siridan himoya omili bo'lib xizmat qiladi. Juftlashgan allel genlari, agar ular tabiatda retsessiv bo'lsa, mutatsiyalarning fenotipik ko'rinishini oldini oladi.
Hayotiy makromolekulalarni kodlovchi genlarni ekstrakopiya qilish hodisasi gen mutatsiyalarining zararli ta'sirini kamaytirishga yordam beradi. Masalan, rRNK, tRNK, giston oqsillari genlari, ularsiz har qanday hujayraning hayotiy faoliyati mumkin emas.
Sanab o'tilgan mexanizmlar evolyutsiya jarayonida tanlangan genlarning saqlanishiga va shu bilan birga populyatsiya genofondida allellarning to'planishiga, irsiy o'zgaruvchanlik zaxirasini shakllantirishga yordam beradi.
94. Genomik mutatsiyalar: poliploidiya, gaploidiya, geteroploidiya. Ularning paydo bo'lish mexanizmlari.
Genomik mutatsiyalar xromosomalar sonining o'zgarishi bilan bog'liq. Genomik mutatsiyalar o'z ichiga oladi geteroploidiya, gaploid va poliploidiya.
Poliploidiya- meiozning buzilishi natijasida butun xromosoma to'plamlarini qo'shish orqali xromosomalarning diploid sonining ko'payishi.
Poliploid shakllarda xromosomalar sonining ko'payishi kuzatiladi, bu gaploid to'plamning karrali: 3n - triploid; 4n - tetraploid, 5n - pentaploid va boshqalar.
Poliploid shakllari diploidlardan fenotipik jihatdan farq qiladi: xromosomalar sonining o'zgarishi bilan birga irsiy xususiyatlar ham o'zgaradi. Poliploidlarda hujayralar odatda katta bo'ladi; ba'zan o'simliklar ulkan bo'ladi.
Bitta genom xromosomalarining ko'payishi natijasida hosil bo'lgan shakllar avtoploid deb ataladi. Biroq, poliploidiyaning yana bir shakli ham ma'lum - ikki xil genomning xromosomalari soni ko'payadigan alloploidiya.
Somatik hujayralar xromosomalari sonining diploid bilan solishtirganda bir necha marta kamayishi deyiladi. gaploid... Gaploid organizmlar tabiiy muhitda asosan o'simliklar, shu jumladan yuqori o'simliklar (do'ppi, bug'doy, makkajo'xori) orasida uchraydi. Bunday organizmlarning hujayralari har bir gomologik juftlikning bitta xromosomasiga ega, shuning uchun fenotipda barcha retsessiv allellar paydo bo'ladi. Bu gaploidlarning hayotiy qobiliyatining pasayishini tushuntiradi.
Geteroploidiya... Mitoz va meiozning buzilishi natijasida xromosomalar soni o'zgarishi va gaploid to'plamining ko'paytirilmasligi mumkin. Xromosomalarning birortasi juft bo'lish o'rniga uch karra bo'lib chiqishi hodisasi shunday nom oldi. trisomiyalar... Agar bitta xromosomada trisomiya kuzatilsa, unda bunday organizm trisomik deb ataladi va uning xromosoma to'plami 2n + 1 ga teng. Trisomiya xromosomalarning har qandayida va hatto bir nechtasida bo'lishi mumkin. Ikki marta trisomiya bilan u 2n + 2, uchlik - 2n + 3 va hokazo xromosomalar to'plamiga ega.
Qarama-qarshi hodisa trisomiyalar, ya'ni. diploid to'plamdagi juftlikdan xromosomalardan birining yo'qolishi deyiladi monosomiya, organizm monosomikdir; uning genotipik formulasi 2n-1. Ikki xil xromosoma bo'lmasa, organizm 2n-2 genotipik formulaga ega bo'lgan qo'sh monosomal va boshqalar.
Aytilganlardan ma'lum bo'ladiki anevloidiya, ya'ni. xromosomalarning normal sonining buzilishi, strukturaning o'zgarishiga va organizmning hayotiyligining pasayishiga olib keladi. Buzilish qanchalik katta bo'lsa, yashash qobiliyati shunchalik past bo'ladi. Odamlarda xromosomalarning muvozanatli to'plamining buzilishi xromosoma kasalliklari deb ataladigan og'riqli holatlarga olib keladi.
Vujudga kelish mexanizmi genomik mutatsiyalar meiozda xromosomalarning normal ajralishining buzilishi patologiyasi bilan bog'liq bo'lib, buning natijasida g'ayritabiiy gametalar hosil bo'ladi, bu mutatsiyalarga olib keladi. Tanadagi o'zgarishlar genetik jihatdan o'xshash bo'lmagan hujayralar mavjudligi bilan bog'liq.
95. Inson irsiyatini o'rganish usullari. Genealogik va egizak usullar, ularning tibbiyot uchun ahamiyati.
Inson irsiyatini o'rganishning asosiy usullari quyidagilardir genealogik, egizak, populyatsiya-statistik, dermatoglifika usuli, sitogenetik, biokimyoviy, somatik hujayra genetikasi usuli, modellashtirish usuli
Genealogik usul. Bu usul naslchilikni tuzish va tahlil qilishga asoslangan. Naslchilik - bu oila a'zolari o'rtasidagi munosabatlarni aks ettiruvchi diagramma. Naslchilikni tahlil qilib, ular oilaviy aloqada bo'lgan odamlarning avlodlarida har qanday normal yoki (ko'pincha) patologik belgilarni o'rganadilar.
Genealogik usullar belgining irsiy yoki noirsiyligini, dominantligi yoki retsessivligini aniqlash, xromosoma xaritasini tuzish, jinslar bogʻlanishi, mutatsiya jarayonini oʻrganish uchun qoʻllaniladi. Qoida tariqasida, genealogik usul tibbiy genetik maslahatda xulosalar uchun asos bo'ladi.
Naslchilikni tuzishda standart belgilar qo'llaniladi. Tadqiqotni boshlagan odam probanddir. Er-xotinning avlodi aka-uka, opa-singillar aka-uka, amakivachchalar qarindoshlar aka-uka va boshqalar deb ataladi. Onasi umumiy (lekin otalari har xil) boʻlgan avlodlar qarindoshlar, otalari umumiy (lekin onalari har xil) boʻlgan avlodlar qarindoshlar deyiladi; agar oilada turli nikohdan bo‘lgan farzandlar bo‘lsa, bundan tashqari, ularning umumiy ajdodlari bo‘lmasa (masalan, onaning birinchi nikohidan farzand va otaning birinchi nikohidan bo‘lgan bola), u holda ular yarim ko‘ngilli deb ataladi.
Genealogik usul yordamida o'rganilayotgan belgining irsiy shartliligi, shuningdek, meros turini aniqlash mumkin. Bir nechta belgilar bo'yicha nasl-nasabni tahlil qilganda, ularning merosxo'rligining bog'liqligini aniqlash mumkin, bu xromosoma xaritalarini tuzishda qo'llaniladi. Bu usul mutatsiya jarayonining intensivligini o'rganish, allelning ekspressivligi va penetranligini baholash imkonini beradi.
Ikkilik usuli... Bu bitta va qo'sh egizaklarning juftliklarida belgilarning irsiylanish qonuniyatlarini o'rganishdan iborat. Egizaklar ikki yoki undan ortiq bola bo'lib, deyarli bir vaqtning o'zida bir ona tomonidan homilador va tug'iladi. Bir xil va birodar egizaklarni farqlang.
Bir xil (monozigot, bir xil) egizaklar zigota parchalanishining dastlabki bosqichlarida, ajralish paytida ikki yoki to'rtta blastomera to'laqonli organizmga aylanish qobiliyatini saqlab qolganda paydo bo'ladi. Zigota mitoz yo'li bilan bo'linganligi sababli, bir xil egizaklarning genotiplari, hech bo'lmaganda, boshida butunlay bir xil bo'ladi. Bir xil egizaklar har doim bir jinsli bo'lib, intrauterin rivojlanish davrida ular bitta platsentaga ega.
Ikki yoki undan ortiq bir vaqtning o'zida pishgan tuxum urug'lantirilganda turli xil tuxumlar (dizigotik, bir xil bo'lmagan) paydo bo'ladi. Shunday qilib, ular umumiy genlarning taxminan 50% ni bo'lishadi. Boshqacha qilib aytganda, ular genetik konstitutsiyada oddiy aka-uka va opa-singillarga o'xshaydi va bir jinsli yoki qarama-qarshi jins bo'lishi mumkin.
Xuddi shu muhitda tarbiyalangan bir xil va birodar egizaklarni solishtirganda, belgilarning rivojlanishida genlarning roli haqida xulosa chiqarish mumkin.
Egizaklar usuli sizga xususiyatlarning irsiyligi to'g'risida asosli xulosalar chiqarishga imkon beradi: irsiyat, atrof-muhit va tasodifiy omillarning shaxsning ayrim xususiyatlarini aniqlashdagi roli
Irsiy patologiyaning oldini olish va tashxislash
Hozirgi vaqtda irsiy patologiyaning oldini olish to'rt darajada amalga oshiriladi: 1) pregametik; 2) prezigotik; 3) prenatal; 4) neonatal.
1.) Pregametik daraja
Amalga oshirildi; bajarildi:
1. Ishlab chiqarishni sanitariya nazorati - mutagenlarning organizmga ta'sirini istisno qilish.
2. Tug'ish yoshidagi ayollarni xavfli ishlarda ishlashdan ozod qilish.
3.Ro'yxatlarni yaratish irsiy kasalliklar ma'lum bir narsada keng tarqalgan
def bilan hudud. tez-tez.
2. Prezigotik daraja
Profilaktikaning ushbu darajasining eng muhim elementi aholining tibbiy genetik maslahati (MGK) bo'lib, u oilaga tekshiruv patologiyasi bo'lgan bola tug'ilish xavfi darajasi haqida ma'lumot beradi va tug'ish to'g'risida to'g'ri qaror qabul qilishga yordam beradi.
Prenatal daraja
Bu prenatal (prenatal) diagnostikani o'tkazishdan iborat.
Prenatal diagnostika Homiladagi irsiy patologiyani aniqlash va ushbu homiladorlikni to'xtatish uchun amalga oshiriladigan chora-tadbirlar majmui. Prenatal diagnostika usullariga quyidagilar kiradi:
1. Ultrasonik skanerlash (USS).
2. Fetoskopiya- optik tizim bilan jihozlangan elastik prob orqali bachadon bo'shlig'ida homilani vizual kuzatish usuli.
3... Chorion biopsiyasi... Usul xorionik villi olish, hujayralarni etishtirish va ularni sitogenetik, biokimyoviy va molekulyar genetik usullar yordamida tekshirishga asoslangan.
4. Amniyosentez- qorin devori orqali amniotik suyuqlikni ponksiyon qilish va olish
amniotik suyuqlik. U tekshirilishi mumkin bo'lgan xomilalik hujayralarni o'z ichiga oladi
homilaning taxmin qilingan patologiyasiga qarab sitogenetik yoki biokimyoviy.
5. Kordosentez- kindik ichak tomirlarini ponksiyon qilish va homila qonini olish. Xomilaning limfotsitlari
yetishtiriladi va sinovdan o'tkaziladi.
4.Neonatal daraja
To'rtinchi bosqichda yangi tug'ilgan chaqaloqlar klinikadan oldingi bosqichda autosomal retsessiv metabolik kasalliklarni aniqlash uchun skriningdan o'tkaziladi, bunda o'z vaqtida boshlangan davolanish normal aqliy va ruhiy holatni ta'minlashga imkon beradi. jismoniy rivojlanish bolalar.
Irsiy kasalliklarni davolash tamoyillari
Davolashning quyidagi turlari mavjud.
1. Semptomatik(kasallik belgilariga ta'siri).
2. Patogenetik(kasallikning rivojlanish mexanizmlariga ta'siri).
Semptomatik va patogenetik davolash kasallikning sabablarini bartaraf etmaydi, chunki bartaraf etmaydi
genetik nuqson.
Semptomatik va patogenetik davolashda quyidagi usullardan foydalanish mumkin.
· Tuzatish jarrohlik usullari bilan nuqsonlar (sindaktiliya, polidaktiliya,
yuqori labning yopilmasligi ...
O'rnini bosuvchi terapiya, uning ma'nosi tanaga kiritishdir
etishmayotgan yoki etarli bo'lmagan biokimyoviy substratlar.
· Metabolizm induksiyasi- sintezni kuchaytiruvchi moddalarni organizmga kiritish
ba'zi fermentlar va shuning uchun jarayonlarni tezlashtiradi.
· Metabolizmni inhibe qilish- bog'lovchi va olib tashlaydigan dorilarni tanaga kiritish
anormal metabolik mahsulotlar.
· Dietoterapiya ( tibbiy ovqatlanish) - dietadan moddalarni chiqarib tashlash
organizm tomonidan so'rilmaydi.
Istiqbollar: Yaqin kelajakda genetika tez rivojlanadi, garchi u hali ham bizning kunlarimizda.
ekinlarda juda keng tarqalgan (naslchilik, klonlash),
tibbiyot (tibbiy genetika, mikroorganizmlar genetikasi). Kelajakda olimlar umid qilmoqda
nuqsonli genlarni yo'q qilish va yuqadigan kasalliklarni yo'q qilish uchun genetikadan foydalaning
meros orqali, saraton, virusli kabi jiddiy kasalliklarni davolay olish
infektsiyalar.
Radiogenetik ta'sirni zamonaviy baholashning barcha kamchiliklari bilan, atrof-muhitdagi radioaktiv fonning nazoratsiz o'sishi taqdirda insoniyatni kutayotgan genetik oqibatlarning jiddiyligiga shubha yo'q. Atom va vodorod qurollarini keyingi sinovdan o'tkazish xavfi aniq.
Shu bilan birga, genetika va naslchilikda atom energiyasidan foydalanish o'simliklar, hayvonlar va mikroorganizmlarning irsiyatini nazorat qilishning yangi usullarini yaratish, organizmlarning genetik moslashuv jarayonlarini yaxshiroq tushunish imkonini beradi. Odamlarning parvozlari bilan bog'liq holda bo'sh joy kosmik reaksiyaning tirik organizmlarga ta'sirini o'rganish zarur bo'ladi.
98. Odamning xromosoma anomaliyalarini diagnostika qilishning sitogenetik usuli. Amniyosentez. Odam xromosomalarining kariotipi va idiogrammasi. Biokimyoviy usul.
Sitogenetik usul mikroskop yordamida xromosomalarni o'rganishdan iborat. Ko'pincha tadqiqot ob'ekti mitotik (metafaza), kamroq tez-tez meiotik (profaza va metafaza) xromosomalardir. Ayrim shaxslarning karyotiplarini o'rganishda sitogenetik usullar qo'llaniladi
Rivojlanayotgan intrauterin organizmning materialini olish amalga oshiriladi turli yo'llar bilan... Ulardan biri amniyosentez, uning yordamida homiladorlikning 15-16 xaftasida homila va uning terisi va shilliq pardalari hujayralarining chiqindilari bo'lgan amniotik suyuqlik olinadi.
Amniyosentez paytida olingan material biokimyoviy, sitogenetik va molekulyar kimyoviy tadqiqotlar uchun ishlatiladi. Sitogenetik usullar homilaning jinsini aniqlaydi va xromosoma va genomik mutatsiyalarni aniqlaydi. Amniotik suyuqlik va xomilalik hujayralarni biokimyoviy usullar yordamida o'rganish genlarning oqsil mahsulotlaridagi nuqsonni aniqlashga imkon beradi, ammo genomning tarkibiy yoki tartibga soluvchi qismida mutatsiyalarning lokalizatsiyasini aniqlashga imkon bermaydi. DNK problaridan foydalanish irsiy kasalliklarni aniqlash va homilaning irsiy materialiga zararni aniq lokalizatsiya qilishda muhim rol o'ynaydi.
Hozirgi vaqtda amniyosentez yordamida barcha xromosoma anomaliyalari, 60 dan ortiq irsiy metabolik kasalliklar, ona va homilaning eritrotsitlar antigenlari uchun mos kelmasligi aniqlanadi.
Hujayra xromosomalarining soni, hajmi va shakli bilan tavsiflangan diploid to'plami deyiladi. karyotip... Oddiy inson karyotipi 46 xromosoma yoki 23 juftni o'z ichiga oladi: ulardan 22 juft autosoma va bir juft jinsiy xromosomadir.
Karyotipni tashkil etuvchi xromosomalarning murakkab kompleksini tushunishni osonlashtirish uchun ular shaklda joylashtirilgan. idiogrammalar... V idiogramma xromosomalar juft-juft bo'lib kattaligining kamayishi tartibida joylashtirilgan, jinsiy xromosomalar bundan mustasno. Eng katta juftlikka 1-son, eng kichigi - 22-son berildi. Xromosomalarni faqat o'lchamiga qarab aniqlash katta qiyinchiliklarga duch keladi: bir qator xromosomalar o'xshash o'lchamlarga ega. Biroq, ichida yaqin vaqtlar turli xil bo'yoqlardan foydalangan holda, inson xromosomalarining uzunligi bo'ylab maxsus usullar bilan bo'yalgan va bo'yalmagan chiziqlarga aniq farqlanishi o'rnatildi. Xromosomalarni aniq farqlash qobiliyati tibbiy genetika uchun katta ahamiyatga ega, chunki u odamning karyotipidagi buzilishlarning xarakterini aniq aniqlash imkonini beradi.
Biokimyoviy usul
99. Inson kariotipi va idiogrammasi. Inson karyotipining xarakteristikalari normaldir
va patologiya.
Karyotip- xromosomalarning to'liq to'plamining belgilar to'plami (soni, hajmi, shakli va boshqalar);
ma'lum bir biologik turning (turlarning karyotipi), ma'lum bir organizmning hujayralariga xosdir
(individual karyotip) yoki hujayralar chizig'i (klon).
Karyotipni aniqlash uchun mikrografiya yoki bo'linuvchi hujayralar mikroskopi bilan xromosomalarning eskizi qo'llaniladi.
Har bir insonda 46 ta xromosoma bor, ulardan ikkitasi jinsdir. Ayolda ikkita X xromosoma mavjud
(karyotip: 46, XX), erkaklarda bitta X xromosoma, ikkinchisi Y (karyotip: 46, XY) mavjud. O'qish
Karyotip sitogenetik deb ataladigan usul yordamida amalga oshiriladi.
Idiogramma- organizm xromosomalarining haploid to'plamining sxematik tasviri, qaysi
hajmiga qarab bir qatorda, kattaligining kamayishiga qarab juft-juft bo'lib joylashtirilgan. Jinsiy xromosomalar bundan mustasno, ayniqsa ajralib turadi.
Eng keng tarqalgan xromosoma anomaliyalariga misollar.
Daun sindromi - bu 21-juft xromosomalarda trisomiya.
Edvards sindromi - 18-juft xromosomalarda trisomiya.
Patau sindromi - 13-juft xromosomalarda trisomiya.
Klaynfelter sindromi o'g'il bolalarda X xromosoma polisomiyasidir.
100. Genetikaning tibbiyot uchun ahamiyati. Odam irsiyatini o'rganishning sitogenetik, biokimyoviy, populyatsiya-statistik usullari.
Inson hayotida genetikaning o'rni juda katta. U tibbiy genetik maslahat yordamida amalga oshiriladi. Tibbiy genetik maslahat insoniyatni irsiy (genetik) kasalliklar bilan bog'liq azoblardan qutqarish uchun mo'ljallangan. Tibbiy genetik maslahatning asosiy maqsadi ma'lum bir kasallikning rivojlanishida genotipning rolini aniqlash va kasal nasl tug'ilish xavfini bashorat qilishdir. Tibbiy-genetik konsultatsiyalarda nikoh yoki naslning genetik foydaliligi prognozi bo'yicha berilgan tavsiyalar ularni ixtiyoriy ravishda tegishli qaror qabul qiladigan maslahatchilar tomonidan hisobga olinishini ta'minlashga qaratilgan.
Sitogenetik (karyotipik) usul. Sitogenetik usul mikroskop yordamida xromosomalarni o'rganishdan iborat. Ko'pincha tadqiqot ob'ekti mitotik (metafaza), kamroq tez-tez meiotik (profaza va metafaza) xromosomalardir. Bu usul jinsiy xromatinni o'rganish uchun ham qo'llaniladi ( buzoq barra) Ayrim shaxslarning kariotiplarini o'rganishda sitogenetik usullar qo'llaniladi
Sitogenetik usuldan foydalanish nafaqat xromosomalarning normal morfologiyasini va umuman karyotipni o'rganishga, organizmning genetik jinsini aniqlashga, balki, eng muhimi, xromosomalar sonining o'zgarishi bilan bog'liq turli xil xromosoma kasalliklarini aniqlashga imkon beradi. yoki ularning tuzilishining buzilishi. Bundan tashqari, bu usul mutagenez jarayonlarini xromosomalar va karyotip darajasida o'rganish imkonini beradi. Xromosoma kasalliklarini prenatal diagnostika qilish uchun tibbiy-genetik maslahatlarda foydalanish homiladorlikni o'z vaqtida to'xtatish orqali rivojlanishning qo'pol buzilishi bo'lgan nasllarning paydo bo'lishining oldini olishga imkon beradi.
Biokimyoviy usul qon yoki siydikda fermentlarning faolligini yoki ba'zi metabolik mahsulotlarning tarkibini aniqlashdan iborat. Yordamida bu usul ular genotipdagi allel genlarning noqulay kombinatsiyasi, ko'pincha homozigot holatida retsessiv allellar mavjudligi sababli metabolik kasalliklarni aniqlaydi. Bunday irsiy kasalliklarni o'z vaqtida tashxislash bilan profilaktika choralari jiddiy rivojlanish buzilishlaridan qochish imkonini beradi.
Aholi-statistik usul. Bu usul ma'lum bir populyatsiya guruhida yoki yaqin qarindosh bo'lgan nikohlarda ma'lum bir fenotipga ega bo'lgan shaxslarning tug'ilish ehtimolini baholashga imkon beradi; retsessiv allellarning geterozigota holatida tashish chastotasini hisoblang. Usul Hardy - Weinberg qonuniga asoslanadi. Xardi-Vaynberg qonuni Populyatsiya genetikasi qonuni. Qonunda shunday deyilgan: "Ideal populyatsiyada genlar va genotiplarning chastotalari avloddan-avlodga o'zgarmas bo'lib qoladi".
Inson populyatsiyalarining asosiy belgilari: umumiy hudud va erkin nikoh imkoniyati. Yakkalanish omillari, ya'ni turmush o'rtoqlarni tanlash erkinligini cheklash, inson nafaqat geografik, balki diniy va ijtimoiy to'siqlarga ham ega bo'lishi mumkin.
Bundan tashqari, bu usul mutatsiya jarayonini, odamlarda fenotipik polimorfizmning normal belgilarga ko'ra shakllanishida, shuningdek, kasalliklarning, ayniqsa, irsiy moyilligi bo'lgan kasalliklarning paydo bo'lishida irsiyat va muhitning rolini o'rganish imkonini beradi. Antropogenezda, xususan, irq shakllanishida irsiy omillarning ahamiyatini aniqlash uchun populyatsiya-statistik usuldan foydalaniladi.
101. Xromosomalarning struktura aberratsiyasi (aberratsiyasi). Genetik materialning o'zgarishiga asoslangan tasnif. Biologiya va tibbiyot uchun ahamiyati.
Xromosoma aberatsiyasi xromosomalarning qayta joylashishi natijasida yuzaga keladi. Ular xromosomaning yorilishi oqibati bo'lib, keyinchalik qayta birlashtirilgan bo'laklarning paydo bo'lishiga olib keladi, ammo xromosomaning normal tuzilishi tiklanmaydi. Xromosoma aberatsiyasining 4 ta asosiy turi mavjud: tanqislik, ikki barobar, inversiya, translokatsiyalar, o'chirish- xromosoma tomonidan ma'lum bir hududning yo'qolishi, keyinchalik odatda yo'q qilinadi
Kamchiliklar ma'lum bir saytning xromosomasini yo'qotish natijasida paydo bo'ladi. Xromosomaning o'rta qismidagi nuqsonlar deletsiya deb ataladi. Xromosomaning muhim qismini yo'qotish tanani o'limga olib keladi, ahamiyatsiz joylarni yo'qotish irsiy xususiyatlarning o'zgarishiga olib keladi. Shunday qilib. Makkajo'xori tarkibida xromosomalardan biri yetishmasa, uning ko'chatlarida xlorofill yo'q bo'ladi.
Ikki marta ko'paytirish xromosomaning qo'shimcha, dublikat qismini kiritish bilan bog'liq. Bu ham yangi belgilar paydo bo'lishiga olib keladi. Shunday qilib, Drosophilada chiziqli ko'zlar uchun gen xromosomalardan birining bo'limining ko'payishi bilan bog'liq.
Inversiyalar xromosoma buzilganda va ajratilgan joy 180 gradusga aylantirilganda kuzatiladi. Agar yorilish bir joyda sodir bo'lsa, ajratilgan bo'lak xromosomaga qarama-qarshi uchi bilan biriktiriladi, lekin agar ikkita joyda bo'lsa, o'rta bo'lak burilish joylariga, lekin turli uchlari bilan biriktiriladi. Darvinning fikricha, turlarning evolyutsiyasida inversiya muhim rol o'ynaydi.
Translokatsiyalar bir juftdan xromosoma bo'limi gomologik bo'lmagan xromosomaga biriktirilganda paydo bo'ladi, ya'ni. boshqa juftlikdagi xromosoma. Translokatsiya xromosomalardan birining bo'limlari odamlarda ma'lum; bu Daun kasalligining sababi bo'lishi mumkin. Xromosomalarning katta qismlarini o'z ichiga olgan translokatsiyalarning aksariyati organizmni yaroqsiz holga keltiradi.
Xromosoma mutatsiyalari ba'zi genlarning dozasini o'zgartirish, bog'lanish guruhlari o'rtasida genlarning qayta taqsimlanishiga olib kelishi, ularning bog'lanish guruhidagi lokalizatsiyasini o'zgartirish. Bu bilan ular organizm hujayralarining gen muvozanatini buzadilar, buning natijasida shaxsning somatik rivojlanishida og'ishlar mavjud. Odatda, o'zgarishlar bir nechta organ tizimlariga ta'sir qiladi.
Xromosoma aberatsiyasi tibbiyotda katta ahamiyatga ega. Da xromosoma aberatsiyasi, umumiy jismoniy va kechikish bor aqliy rivojlanish... Xromosoma kasalliklari ko'plab tug'ma nuqsonlarning kombinatsiyasi bilan tavsiflanadi. Bunday nuqson Daun sindromining namoyon bo'lishi bo'lib, u 21-xromosomaning uzun qo'lining kichik segmentida trisomiya holatida kuzatiladi. Yig'layotgan sindromning rasmi 5-xromosomaning qisqa qo'lining bir qismini yo'qotish bilan rivojlanadi. Odamlarda miya, mushak-skelet, yurak-qon tomir va genitouriya tizimlarining malformatsiyasi ko'pincha kuzatiladi.
102. Tur tushunchasi, turlanish haqidagi zamonaviy qarashlar. Ko'rish mezonlari.
Ko'rinish Turlarning mezonlari bo'yicha bir-biriga o'xshash bo'lgan shaxslar to'plami, ular shunday darajada
tabiiy ravishda chatishadi va unumdor nasl beradi.
Fertil nasl- o'zi ko'paytira oladigan narsa. Bepusht nasllarga xachir (eshak va otning duragaylari) misol bo'la oladi, u bepushtdir.
Ko'rish mezonlari- bu 2 ta organizmning bir turga mansubligini yoki har xilligini aniqlash uchun solishtiriladigan belgilar.
· Morfologik - ichki va tashqi tuzilishi.
· Fiziologik va biokimyoviy - organlar va hujayralar qanday ishlaydi.
· Xulq-atvor - xatti-harakatlar, ayniqsa ko'payish davrida.
Ekologik - hayot uchun zarur bo'lgan ekologik omillar majmui
turlari (harorat, namlik, oziq-ovqat, raqobatchilar va boshqalar)
Geografik - hudud (tarqatish maydoni), ya'ni. bu tur yashaydigan hudud.
· Genetik-reproduktiv - xromosomalarning soni va tuzilishi bir xil bo'lib, bu organizmlarga unumdor nasl berish imkonini beradi.
Ko'rish mezonlari nisbiy, ya'ni. turlarni baholash uchun bitta mezondan foydalanish mumkin emas. Masalan, qardosh turlari (bezgak chivinlarida, kalamushlarda va boshqalar) mavjud. Ular morfologik jihatdan bir-biridan farq qilmaydi, lekin ular turli xil miqdordagi xromosomalarga ega va shuning uchun nasl bermaydilar.
103. Aholi. Uning ekologik va genetik xususiyatlari va turlanishdagi roli.
Aholi- bir turdagi individlarning o'z-o'zidan ko'payadigan, boshqa shunga o'xshash guruhlardan ozmi-ko'pmi ajratilgan, ma'lum bir hududda uzoq avlodlar silsilasi davomida yashaydigan, o'ziga xos genetik tizimini shakllantiradigan va o'zining ekologik o'rnini shakllantiradigan minimal guruhi.
Aholining ekologik ko'rsatkichlari.
soni- populyatsiyadagi individlarning umumiy soni. Bu qiymat o'zgaruvchanlikning keng doirasi bilan tavsiflanadi, lekin u ma'lum chegaralardan past bo'lishi mumkin emas.
Zichlik- maydon yoki hajm birligidagi individlar soni. Raqamlarning ko'payishi bilan aholi zichligi, qoida tariqasida, ortadi
Fazoviy tuzilma aholi bosib olingan hududda individlarning tarqalishining o'ziga xos xususiyatlari bilan tavsiflanadi. U yashash joyining xususiyatlari va turning biologik xususiyatlari bilan belgilanadi.
Gender tuzilishi aholida erkak va ayolning ma'lum nisbatini aks ettiradi.
Yosh tuzilishi umr ko'rish davomiyligi, balog'atga etish vaqti, avlodlar soniga qarab populyatsiyalarda turli yosh guruhlari nisbatini aks ettiradi.
Populyatsiyaning genetik ko'rsatkichlari... Genetik jihatdan populyatsiya genofondi bilan tavsiflanadi. U ma'lum bir populyatsiyadagi organizmlarning genotiplarini tashkil etuvchi allellar to'plami bilan ifodalanadi.
Populyatsiyalarni tavsiflashda yoki ularni bir-biri bilan solishtirishda bir qator genetik xususiyatlardan foydalaniladi. Polimorfizm... Agar ikki yoki undan ortiq allellar topilgan bo'lsa, populyatsiya ma'lum bir joyda polimorf deyiladi. Agar lokus bitta allel bilan ifodalangan bo'lsa, u monomorfizm haqida gapiradi. Ko'pgina lokuslarni o'rganib, ular orasida polimorflarning ulushini aniqlash mumkin, ya'ni. populyatsiyaning genetik xilma-xilligining ko'rsatkichi bo'lgan polimorfizm darajasini baholash.
Geterozigotalik... Populyatsiyaning muhim genetik xarakteristikasi geterozigotalikdir - populyatsiyadagi geterozigotalarning chastotasi. Shuningdek, u genetik xilma-xillikni aks ettiradi.
Inbreeding koeffitsienti... Ushbu koeffitsient populyatsiyada bir-biriga yaqin bo'lgan xochlarning tarqalishini baholash uchun ishlatiladi.
Genlar assotsiatsiyasi... Turli genlarning allel chastotalari bir-biriga bog'liq bo'lishi mumkin, bu assotsiatsiya koeffitsientlari bilan tavsiflanadi.
Genetik masofalar. Turli xil populyatsiyalar allel chastotalarida farqlanadi. Ushbu farqlarni aniqlash uchun genetik masofalar deb ataladigan ko'rsatkichlar taklif qilingan.
Aholi- elementar evolyutsion tuzilish. Har qanday tur oralig'ida individlar notekis taqsimlangan. Jismoniy shaxslarning zich kontsentratsiyasi joylari ular ko'p bo'lmagan yoki yo'q bo'lgan bo'shliqlar bilan kesishadi. Natijada, tasodifiy erkin kesishish (panmiksiya) muntazam ravishda sodir bo'ladigan ko'proq yoki kamroq izolyatsiya qilingan populyatsiyalar paydo bo'ladi. Boshqa populyatsiyalar bilan chatishtirish juda kam va tartibsiz. Panmiksiya tufayli har bir populyatsiyada boshqa populyatsiyalardan farq qiladigan xarakterli genofond hosil bo'ladi. Aynan populyatsiya evolyutsiya jarayonining elementar birligi sifatida tan olinishi kerak.
Populyatsiyalarning roli katta, chunki deyarli barcha mutatsiyalar uning ichida sodir bo'ladi. Bu mutatsiyalar, birinchi navbatda, populyatsiyalar va genofondning izolyatsiyasi bilan bog'liq bo'lib, ular bir-biridan ajratilganligi sababli farqlanadi. Evolyutsiya uchun material populyatsiyada boshlanib, turning shakllanishi bilan tugaydigan mutatsion o'zgaruvchanlikdir.